Introduction
2012년 개정된 양돈 NRC (2012)에 약 120종의 사용 가능한 원료사료들이 수록되어 있으나 국내에서 활용되고 있는 원료사료들은 기호성 및 성장에 효과적인 옥수수 및 대두박를 주로 이용하고 있다(Park et al., 2016). 맥류 원료사료인 보리는 영양소가 균형적으로 포함되어 있는 우
수한 원료사료 중 하나이다. 특히, 쌀보리는 껍질이 잘 분리되어 식용으로 사용되는 식량자원이나 탄수화물 섭취량의 감소로 재고량이 증가하고 있다(Ahn et al., 2015). 해결방안으로 동물사료원으로 활용하는 방안이 검토되고 있다. 그러나, 보리는 높은 수분함량으로 인해 저장성이 낮고 내배유와 호분층에 존재하는 베타글루칸(β- glucans)은 영양소 소화 및 흡수를 저해하는 것으로 알려져 있다(Li et al., 1996; Ahn et al., 2015). 옥수수에 비해 보리를 급여한 육성돈의 아미노산소화율은 감소시킨다(Sauer et al., 1997). 따라서, 발효와 같은 가공처리를 통해 저장성 향상과 같은 보리의 사료가치의 증진이 필요하다(Ahn et al., 2015).
사료 내 생균제와 같은 미생물의 첨가는 돼지의 생산성을 향상시키며 젖산발효를 통해 장건강에 도움을 주고 궁극적으로 동물의 생산성을 증진시키는 것으로 알려져 있다(Underdahl et al., 1982; Min and Kim, 2002; Altmeyer et al., 2014). 일부 연구에서는 발효된 사료를 직접 급여한 결과 돼지의 성장개선 및 면역기능을 강화시킨다고 하였다(Park et al., 2004; Na et al., 2008; Jeong et al., 2017). 일반적으로 발효에 사용되고 있는 유산균(Lactobacillus sp.)은 약 200종이 보고되어 있으며 Federal Department of Agriculture (FDA)와 European Food Safety Authority (EFSA)의해 안전성이 입증된 유산균은 35종(species)으로 알려져 있다(Euzeby, 1997; London et al., 2014). 이러한 유산균들은 탄수화물이 많은 식물, 사일리지, 우유 그리고 사람 및 동물의 장에서 많이 관찰되며 주변환경이나 사람 또는 동물과 같이 기원이 되는 종에 따라 다양한 유산균이 존재한다(Hammes and Vofel, 1995; Bernardeau et al., 2008). Lee (2012)의 연구에 의하면, 뱀장어 유래 유산균(L. pentosus PL11)은 기존의 L. pentosus JCM1558T와 99.3% 상동성을 나타냈으나 L. pentosus JCM1558T 보다 L. pentosus PL11의 급여가 뱀장어의 면역 및 항산화능 증진에 효과적임을 보고하였다. Ahn et al. (2015)은 쌀보리에는 다수의 미생물들이 분포되어 있으며 분포된 미생물들이 포함된 복합미생물이 접종된 쌀보리에서 정상적인 발효과정이 발생하였다고 보고하였다. 다른 선행연구의 보고에서 들깨 유래 L. plantarum과 L. acidophilus (ATCC4356)를 들깨에 접종 후 발효기간동안 성장속도 및 pH 저하능을 측정한 결과 들깨 유래 L. plantarum이 효과적이다(Bae et al., 2016). 그러므로 유산균은 계통(strain)에 따라 유기산과 같은 기능성물질의 생성능 및 성장속도에서도 차이가 나고 필요로 하는 생육환경도 달라 적용하고자 하는 종에 대한 적합성(species compatibility)이 전제되어야 활용 가능하다(Lee, 2012; Kim and Lee, 2013; London et al., 2014; Ahn et al., 2015).
따라서, 돼지에 적합한 발효균주를 선발하고자 포유자돈 분변으로부터 미생물균총을 조사하고 우점한 미생물의 특성과 발효 쌀보리 내 성분변화에 후보균주가 미치는 영향을 구명하고자 본 연구를 실시하였다.
Materials and Methods
후보균주분리
본 시험에서 쌀보리 발효에 이용될 생균제 후보균주를 선발하기 위해 9 - 10일령 3원교잡종(Landrace × Yorkshire × Duroc) 포유자돈 4마리를 공시하였고 swab법을 이용하여 분변을 채취하였다. 채취한 시료는 0.85% NaCl 용액에서 균질화하고 10배수씩 단계 희석하였다. 시료 내 유산균총 및 일반세균의 분리동정을 위해 MRS agar (BD, Difco, USA)와 Nutrient agar (BD, Difco, USA)에 희석액을 접종하고 각각 혐기 및 호기조건의 37°C 배양기에서 24 h동안 배양하였다. 배양 종료 후 형성된 colony를 계수하였고 각각의 배지에 형성된 단일 colony를 회수한 뒤 동일한 조건에서 분리배양하였다. 분리된 미생물 균종의 동정을 위해 질량분석법(MALDI-TOF, Bruker Microflex TOF mass spectrometer, Bruker Daltonik GmbH, Bremen, Germany)을 이용하였다. 포유자돈의 장내 우점 미생물은 L. plantarum으로 분석되었고 이 균주는 젖산 생성능력이 있는 유산균으로 쌀보리 발효시험에 이용할 후보균주로 선발하였다.
후보균주 내산성 및 유기산 생성능
선발된 균주의 acid tolerance를 확인하기 위해 MRS broth의 pH를 2.5, 3.0 및 7.0으로 각각 조정한 뒤 후보균주 배양액을 1% (v/v) 접종하고 37°C에서 배양하였다. 그리고, 후보균주의 생존성을 확인하기 위해 10배수씩 단계 희석된 용액을 MRS agar에 도말하고 37°C에서 24 h동안 배양한 뒤 colony를 확인하여 내산성을 평가하였다.
후보균주의 배양액을 MRS broth에 1% (v/v) 접종하고 37°C에서 7.5 h동안 배양하였다. 배양시간동안 2.5 h간격으로 배양액을 채취하여 10배수씩 단계 희석 후 MRS agar에 도말하고 37°C에서 24 h동안 배양한 뒤 colony를 확인하여 성장속도를 확인하였다. 0, 2.5, 5.0 및 7.5 h에 채취한 배양액을 원심분리(4°C, 13,000 rpm, 10 min)하고 상층액의 pH를 측정하여 유기산 생성능을 확인하였다. 또한, 후보균주의 내산성과 유기산 생성능을 비교하기 위해 시판 복합생균제(청미락토, ㈜청미바이오)는 대조구로 사용하였다.
후보균주의 쌀보리 발효 및 발효물 특성분석
쌀보리(Hordeum Vulgare L.)의 발효특성 분석을 위해 L. plantarum 배양액(≥ 107 cfu/mL)을 쌀보리에 0.1% 접종하고 밀폐용기에 담아 37°C에서 2일간 배양한 뒤 pH측정하고 10배수씩 단계 희석 후 MRS agar에 도말하고 37°C에서 24 h동안 배양한 뒤 colony를 확인하였다. 균주를 미접종한 쌀보리를 접종한 쌀보리와 동일한 조건에서 배양하여 대조구로 이용하였다.
쌀보리가 발효되는 동안 영양성분의 변화를 측정하기 위해 발효 0, 1, 3, 및 7일차에 발효물의 영양성분을 분석하였다. 분석방법은 AOAC (2005)가 제시하는 방법에 준하여 수분, 조단백질, 조지방, 조섬유, 조회분 함량을 산출하였다. Neutral detergent fiber (NDF)와 acid detergent fiber (ADF)는 Van Soest et al. (1991)의 방법을 사용하여 분석하였다. 에너지수준은 bomb calorimeter (Model 124, Parr Instrument Company, Molin, IL)를 이용하여 제조사에서 제공하는 방법에 의거하여 확인하였다.
통계처리
후보균주 및 발효 쌀보리 특성 구명을 위한 모든 분석은 4반복으로 하였고 반복당 분석횟수는 3회이며 분석치의 평균값을 결과값으로 사용하였다. 수집된 모든 결과는 SAS ver.9.2. (SAS Institute, Carym NC, USA)를 이용해 분산분석을 실시하였고, 처리구간 유의성은 Duncan’s multiple range test을 이용하여 95% 신뢰수준에서 검증하였다.
Results and Discussion
포유자돈의 분변 내 미생물 분포
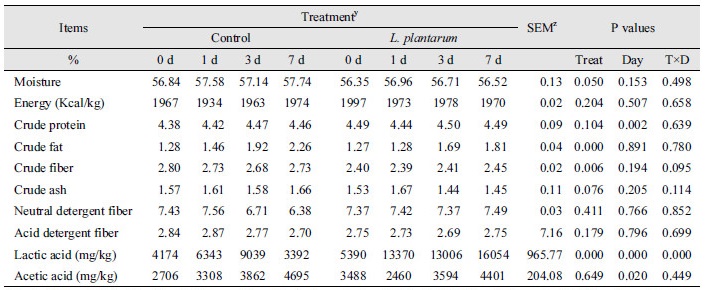
염기서열분석법을 이용하여 포유자돈의 분변으로부터 유래하는 미생물을 동정한 결과는 Fig. 1과 같다. 호기 및 혐기조건에서 분리된 총 206개의 돼지유래 미생물이 동정되었고 가장 우점한 균총은 L. plantarum (n = 133, 64.56%)로 나타났으며 Escherichia coli ETEC H10407 (n = 45, 21.84%)이 두 번째로 많이 존재하였다. Uncultured bacteria는 5.83% (n = 12)의 조성을 보였다. 이외 동정된 균주들은 Enterococcus faecalis (n = 5, 2.43%), Shigella flexneri (n = 4, 1.94%), Bacillus thuringiensis (n = 1, 0.97%), Acidovorax sp. (n = 1, 0.49%), Bacillus subtilis (n = 1, 0.49%), E. coli Xuzhou21 (n = 1, 0.49%), S. epidermidis (n = 1, 0.49%), Staphylococcus warneri (n = 1, 0.49%) 순으로 분석되었다. 분리된 균주 중 젖산을 생산하는 L. plantarum을 쌀보리 발효를 위한 후보균주로 선정하였다.
Fig. 1.
Microbial flora in feces of suckling piglets determined by MALDI-TOF mass spectrometry (n = 206). Lactobacillus plantarum (n = 133), Escherichia coli ETEC H10407 (n = 45), Uncultured bacteria (n = 12), Enterococcus faecalis (n = 5), Shigella flexneri (n = 4), Bacillus thuringiensis (n = 2), Acidovorax sp. (n = 1), Bacillus subtilis (n = 1), Escherichia coli Xuzhou21 (n = 1), Staphylococcus epidermidis (n = 1), Staphylococcus warneri (n = 1).
후보균주의 특성분석
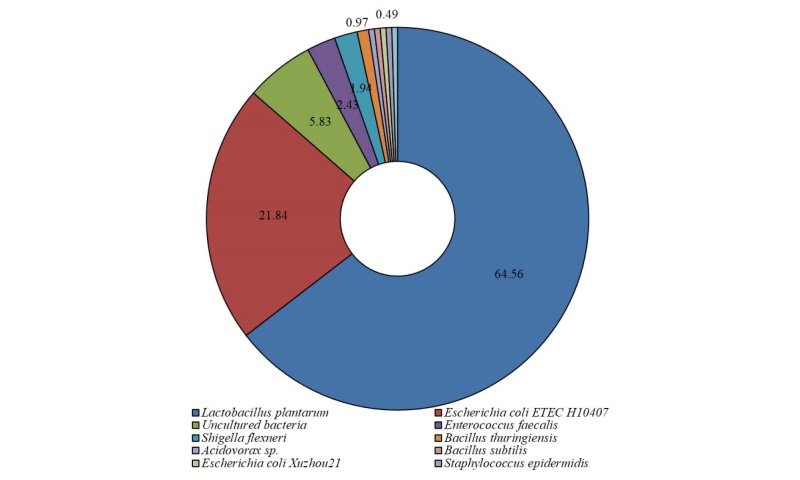
자돈 분변에서 분리된 후보균주의 L. plantarum의 내산성을 확인하고자 pH에 따른 성장을 Table 1에 나타내었다. 생육 시 pH 7.0에서 colony forming unit (cfu)은 5.48 cfu/mL로 pH 3.0 및 2.5의 4.93과 4.56 cfu/mL보다 증가했다(p < 0.05). 강산성인 pH 2.5와 3.0에서 계수된 후보균주은 pH 7보다 각각 16.78%, 10.03% 감소된 것으로 나타났다. 그러나 pH 2.5와 pH 3.0간 후보균주 생육수준은 통계적인 차이를 나타내지 않아 후보균주는 약간의 내산성을 가지는 것으로 판단된다.
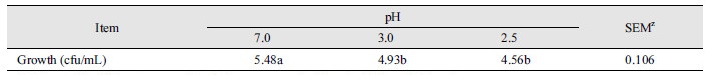
Table 2는 후보균주와 대조구(시판 복합생균제)의 배양시간 별 pH 및 성장변화를 나타내었다. 후보균주와 대조구의 pH 및 성장은 배양 후 0 h에 비해 7.5 h에서 각각 감소 및 증가를 나타냈다(p < 0.05). 이는 배양시험동안 문제가 없으며 정상적으로 생육하여 유기산을 생성하였음을 의미한다. 그러나, 후보균주의 배양시간 별 pH는 대조구에 비해 높게 측정되었다(p < 0.05). 배양시간 5 h 경과 후 후보균주의 성장속도는 대조구에 비해 높았다(p < 0.05). 일반적으로 미생물의 섭취는 구강을 통해 이루어지면 강산성인 위산과 담즙산이 존재하는 위장과 십이지장을 거쳐 장에 도달할 때까지 생존하는 것이 장내 미생물조성에 도움에 된다(Gilliland, 1979; Saarela et al., 2000). 또한, 대조구로 사용된 복합생균제는 단일생균제에 비해 균주들간 상호보완을 통해 균주의 기능을 향상시키는 것으로 알려져 있다(Chapman et al., 2011). 본 연구에서 대조구보다 후보균주는 유기산 생성능은 감소하였으나 성장속도가 빠르며 내산성을 가지는 것으로 측정되어 발효균주로서 활용가능성이 있는 것으로 사료된다.
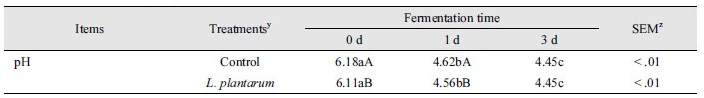
발효 쌀보리 내 pH 변화
후보균주 접종 및 미접종된 쌀보리의 발효동안 pH 변화는 Table 3에 나타냈다. 발효개시일과 발효 1일 및 2일 경과 후 처리구의 pH는 대조구보다 감소하였다(p < 0.05). 시험기간동안 처리구의 pH는 지속적으로 감소하여 발효가 정상적으로 진행됨을 나타냈다(p < 0.05). 그러나 대조구도 시험기간동안 지속적으로 pH가 유의하게 감소하였다(p < 0.05). 본 연구의 이러한 결과는 이전 연구와 일치하였다. Ahn et al. (2015)의 보고에 의하면, 생균제를 미접종한 쌀보리 종실 내 pH는 발효기간에 의한 유의한 감소가 관찰되었다. 쌀보리는 Staphylococcus와 Bacillus속 미생물을 비롯한 다양한 미생물 분포를 가지는 것으로 알려져 있고 이러한 원인은 재배된 토양에서 유래한 미생물들에 의한 것으로 생각되며 자연발효조건하에서 균주 미접종 쌀보리는 유기산 생성이 가능하다(Suh et al., 2010; Ahn et al., 2015). 따라서, 대조구에서의 pH의 감소는 쌀보리에서 유래한 미생물에 기인하는 것으로 사료된다.
미생물을 이용한 발효사료제조는 발효과정에서 유기산이 생성되어 pH가 저하되며 발효초기에 pH저하를 촉진하여 젖산발효를 유도한다(Seal, 1986; Kim et al., 2009). 본 연구에서 포유자돈 유래 L. plantarum을 접종한 발효 쌀보리는 발효초기에 pH 6.11에서 1, 2일차에 pH 4.56 - 4.45로 측정되었다. 이러한 결과는 발효기간동안 젖산을 생산하는 L. plantarum의 성장이 정상적으로 이루어 졌음을 나타낸다. 유산균의 초기 성장에는 당류가 중요한데 당류가 부족한 갈조류의 발효물 내 L. plantarum의 수가 발효 1일차에 0일차보다 오히려 감소됨을 보고하였다(Gupta et al., 2011). 그러므로, 발효사료제조에서 고려할 사항은 젖산생성이 가능한 후보균주의 선택적 이용과 후보균주의 성장에 적절한 기질의 사용이 중요하다. 따라서, 쌀보리는 발효를 위한 기질로서 적합한 것으로 판단된다.
발효 쌀보리 내 화학적 성분 변화
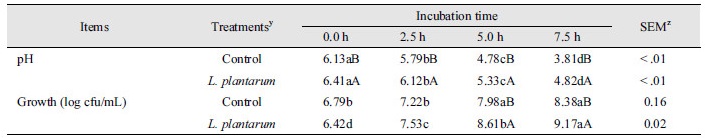
Table 4는 발효기간 별 쌀보리 내 화학적 변화를 나타냈다. 수분함량 및 에너지수준은 발효기간 및 균주의 접종 유무에 따라 영향을 받지 않았다. 조단백질 함량은 처리구간 차이가 없으나 발효기간에 따라 변화를 보였다(p < 0.05). 그러나, 발효기간 중 후보균주 접종구와 대조구의 조단백질 수준은 4.38 - 4.47%로 유의미한 변화로 판단되지 않는다. 발효 쌀보리의 조지방 및 조섬유은 대조구에 비해 접종구에서 감소하였고 조회분은 후보균주의 접종 및 발효기간에 의한 영향은 없었다(p < 0.05). Hemicellulose, cellulose, lignin이 주성분인 NDF와 ADF는 후보균주의 접종 및 발효기간에 의해 차이를 나타내지 않았다. 발효가 사료 내 영양성분에 미치는 영향은 이전부터 보고되어 왔다. 생균제(L. plantarum, Bacillus subtilis) 접종 발효 채종박은 비발효 채종박에 비해 조단백질(58.4 vs. 37.1%), 조지방함량(16.2 vs. 14.8 g/kg)이 증가하며 수분함량(88.1 vs. 93.4%)은 감소하였다(Fazhi et al., 2011). 유산균(L. plantarum)에 의해 발효된 들깨 내 조지방수준은 대조구에 비해 유의하지 않지만 증가했다(72.5 vs.. 71.93 g/100 g; Bae et al., 2016). Jha et al. (2011)의 연구는 in vitro 발효 시 쌀보리의 품종에 따라 건물량(68-80%)에 차이가 있으며 품종에 따라 조단백질수준은 3%의 차이를 나타냈다. 위의 선행연구들은 사료 내 영양성분의 변화가 발효에 의해 야기될 수 있음을 의미한다. 본 연구에서도 조단백질, 조지방, 조섬유수준이 발효 및 접종균주에 의해 차이를 보였으나 언급된 선행연구와 일치하지 않았다. 이러한 차이는 다른 발효기질 및 계통이 다른 유산균 사용에 따른 변화로 판단된다.
발효 쌀보리 내 젖산농도는 발효기간동안 대조구보다 후보균주 접종구에서 통계적으로 증가했다(p < 0.05; Table 4). 아세트산의 발효 쌀보리 내 수준은 발효기간에 의해 유의적으로 증가하였다(p < 0.05). 이러한 결과는 본 연구의 발효 쌀보리 내 pH 변화와 일치한다. 발효는 homo형 및 hetero형으로 나누어 지며 homo형 발효는 젖산만 생성하는 정상발효로 발효효율이 높고 단백질분해를 저해하여 사료의 가치를 높인다(Seal, 1986; Cho et al., 2012). 반면에 hetero형은 젖산 외 초산 및 에탄올 등을 생성하여 homo형에 비해 정상발효과정에서 중요한 pH감소에 효과적이지 않다(Stockes, 1992; Ahn et al., 2015). 따라서, 본 연구에서 후보균주를 이용한 쌀보리의 발효과정은 homo형 발효로 보인다.