
Introduction
커피는 세계적으로 가장 인기있는 음료 중 하나이며, 최근 섭취량 증가로 커피가 체내에 미치는 영향에 대한 많은 연구들이 진행되어 왔다. 그 결과 커피가 인간의 건강에 미치는 영향이 적지 않음이 확인되었다(Hernan et al., 2002; Inoue et al., 2005; Nakayama and Oishi, 2013; Zelber-Sagi et al., 2015; Tran et al., 2019). 예를 들어, 하루에 커피 5잔을 섭취하는 사람은 간암 발병률이 대조군에 비해 25%나 감소한다고 알려져 있다(Inoue et al., 2005). 당뇨병이나(van Dam and Feskens, 2002) 퇴행성신경질환(Hernan et al., 2002)을 예방하는 효능이 있음도 보고되었다. 커피 추출액이 생쥐 소장 속에서 유산균의 성장을 촉진시키거나 병원성 세균의 성장을 억제한다고도 밝혀졌다(Nakayama and Oishi, 2013). 이처럼 대부분의 연구들이 커피 섭취가 인체에 미치는 영향을 분석하거나 커피 추출액의 효능 연구에만 집중되고 있다. 반면에 커피의 주요 성분인 카페인의 직접적인 효능에 대한 연구는 부족한 실정이다. 더욱이 카페인이 장내 유익균으로 알려진 유산균의 항균력에 미치는 영향은 아직까지 불분명하다.
인간의 장 속에는 공생세균과 함께 병원성 세균이 공존한다(Ritter et al., 2018). 일반 공생세균으로 분류되는 대장균과는 달리 일부 병원성 대장균이나 살모넬라는 설사나 장염과 같은 장질환을 유발하는 것으로 알려져 있다(Grassl et al., 2010; Stalb et al., 2018). 이와는 달리, Lactobacillus casei나 Bifidobacterium lactis와 같은 유산균들은 장 속에서 병원성 미생물의 성장을 억제하고 몸에 유리한 여러 작용들을 하여 인체에 유익한 기여를 한다고 알려져 있다(Bellavia et al., 2014; Moeinian et al., 2014; Escribano et al., 2018). 이 같은 결과로 인해, 능동적으로 유산균을 섭취하여 질병 발생을 낮추거나, 장내 환경을 개선하려는 프로바이오틱스가 시도되고 있는 실정이다(Thongprayoon et al., 2019). 락토바실러스 균류들의 항균력이 박테리오신이라는 항균물질 분비와 연관이 있다고 알려져 있다(Trivedi et al., 2013). 박테리오신은 유산균이 분비하는 대표적인 항균펩타이드로서 내열성이 크고 상대 세균의 세포막을 손상시키는 작용을 나타내는 것으로 알려져 있다(Sharma and Srivastava, 2014).
이처럼 유산균들은 병원성 미생물에 대해서 높은 항균력을 보일뿐만 아니라 장내 환경개선에도 큰 역할을 한다. 하지만 이와 같은 유산균의 항균력에 직접적인 영향을 미칠 수 있는 음식물 속 물질에 대한 연구는 미미한 실정이다. 따라서 본 연구에서 커피 속 카페인이 유산균의 항균력에 미치는 영향을 평가해 보고자 하였다.
Materials and Methods
균주
Lactobacillus casei (KACC 12413)는 농촌진흥청으로부터 Escherichia coli (KCCM 21052)는 한국미생물보존센터로부터 분양 받아서 사용하였다. 세균 배양 용 배지는 농촌진흥청과 한국미생물보존센터에서 각각 추천하는 배지로, L. casei는 MRS 배지(Difco MRS, BD, USA), E. coli는 LB (Luria-Bertani) 배지(Difco, USA)를 각각 사용하였다. 분양 받은 균주는 1 mL의 액체배지에 넣어 24 시간 배양하였으며, 활성화 된 균주의 용액을 3 mL의 액체배지에 넣어 24시간, 2차 계대 배양하여 실험에 사용하였다(Bellavia et al., 2014).
L. casei배양상등액 분리
L. casei (3 × 106 CFU/mL)를 MRS 배지에 접종한 후, 카페인(0.3 mg/mL)을 처치한 다음 37℃ 배양기에서 24시간 호기 배양하였다. 배양이 끝난 배양액은 얼음 위에서 20분간 정치 후, 13,000 × g, 4℃, 15분 동안 원심분리하여 상등액을 회수하였다. 상등액은 다시 0.2 μL Syringe filter (Advantec, Japan)를 이용하여 여과하여 잔여 세균을 완전히 제거하였다. 여과된 상등액은 활성 변화가 없도록 4℃에 보관하여 실험에 이용하였다(Mirnejad et al., 2013).
항균활성 측정
병원성 세균에 대한 억제능력은 현탁도법으로 확인하였다. 대장균(E. coli)을 LB 배지 이용 밤새 배양하여 예비 배양액을 준비한 다음, 멸균된 새 배지에 1 × 106 CFU/mL로 접종하였다. 이때 카페인을 처치한 L. casei 배양상등액을 대장균 배양액에 10% 농도로 함께 첨가하였다. 처치 후 37℃ 배양기에서 24시간 동안 호기 배양한 다음, microplate reader (Bio-Rad, Hercules, USA)를 이용하여 655 nm의 흡광도로 대장균의 생육을 측정하였다(Gustine et al., 2019).
열 안정성
카페인으로 자극한 L. casei 배양상등액 속 항균물질이 열에 대해서 안정성이 있는지를 알아보기 위해 100℃에서 20분 동안 열처리하였다. 항균시험은 현탁도법으로 수행하였다(Ghadbane et al., 2013).
대장균 세포막 손상
L. casei 배양상등액의 항균효과가 대장균의 세포막 손상과 관련이 있는지를 확인해보고자 하였다. 시험관에 대장균과 L. casei 배양상등액을 처치하고 4시간 동안 반응한 다음, propidium iodide (PI) 염색시약을 첨가하고 섞어준 후 10분간 추가 반응하였다(Mirnejad et al., 2013). 그 후 반응용액 20 uL를 취하여 형광현미경을 통해 관찰하였다. PI는 세포막이 손상된 세균에만 선별적으로 유입되어 핵을 염색한다.
Statistical analysis
모든 실험은 3회 반복으로 행하여 표준오차로 나타내었고 SIGMA-STAT professional statistics software program (Jandel Scientific Software, USA)의 t-test를 이용하여 유의성을 검정하였다(Ghadbane et al., 2013).
Results and Discussion
L. casei항균력에 대한 카페인의 영향
L. casei는 병원성 미생물에 대해 높은 항균 효과를 나타낸다고 알려져 있다(Mirnejad et al., 2013; Bellavia et al., 2014; Moeinian et al., 2014). 따라서 연구자 등은 최근 다량 섭취되고 있는 커피 속 카페인이 L. casei의 항균력에 미치는 영향을 확인해 보고자 하였다. 1 mg/mL 농도의 카페인 자체가 항균력을 나타내기 때문에(Al-Janabi, 2011), 연구자 등은 L. casei에 처치할 카페인 농도를 결정하였다. 이를 위해 L. casei 배양액에 카페인을 농도 별로(0.3 - 1.2 mg/mL) 밤새 처치한 뒤 성장률 변화를 확인하였다. 그러나 1.2 mg/mL 카페인 농도에서도 L. casei의 성장률 감소는 관찰되지 않았다(결과 미제시). 카페인의 직접적인 항균효과를 완전히 배제하기 위해서 가장 낮은 0.3 mg/mL 카페인 농도를 적용하여 이후 실험을 진행하였다. 다음으로 카페인(0.3 mg/mL)을 처치 후 밤새 배양한 L. casei의 배양상등액을 대장균에 처치한 다음 대장균의 성장률 변화를 확인하였다. 기존 연구 결과처럼, 대장균에 대한 L. casei의 높은 항균력이 확인되었다(Fig. 1A). L. casei 배양상등액 처치가 대장균 성장률을 약 24% 정도 감소시켰다. 카페인으로 자극 후 분리한 L. casei 의 배양상등액의 항균력은 약 39%를 나타났다. 이 결과는 카페인 자극이 병원성 대장균에 대한 L. casei 의 항균효과를 높여줬음을 의미한다. 일부 연구에서 고농도의 카페인 자체가 유의한 항균효과를 나타낸다고 알려졌다(Al-Janabi, 2011). 이는 본 실험에서 확인된 카페인 자극 후 증가된 L. casei의 항균력이 카페인의 직접적인 항균 효과에 기인할 수 있음을 보여준다. 이를 확인하기 위해, 대장균에게 카페인을 직접 처치한 후 성장률 변화를 확인하였다. Fig. 1B에서처럼, L. casei를 자극시키기 위해 넣어준 0.3 mg/mL 카페인이 대장균 성장에 영향을 전혀 미치지 않음을 확인하였다. 1.0 mg/mL 이하의 카페인 농도에서도 대장균 성장에 영향이 없었다(결과 미제시). 병원성 대장균에 대해 항균 효과를 나타내는 카페인의 농도가 논문 별로 상이하게 제시되어온 측면이 있지만, 1 mg/mL 이상의 카페인 농도에서 높은 항균력이 나타남은 분명하다(Al-Janabi, 2011). 이 결과를 종합하면, 카페인이 L. casei의 항균물질 생산과정을 촉진시켜 항균력 강화에 관여함을 보여준다.
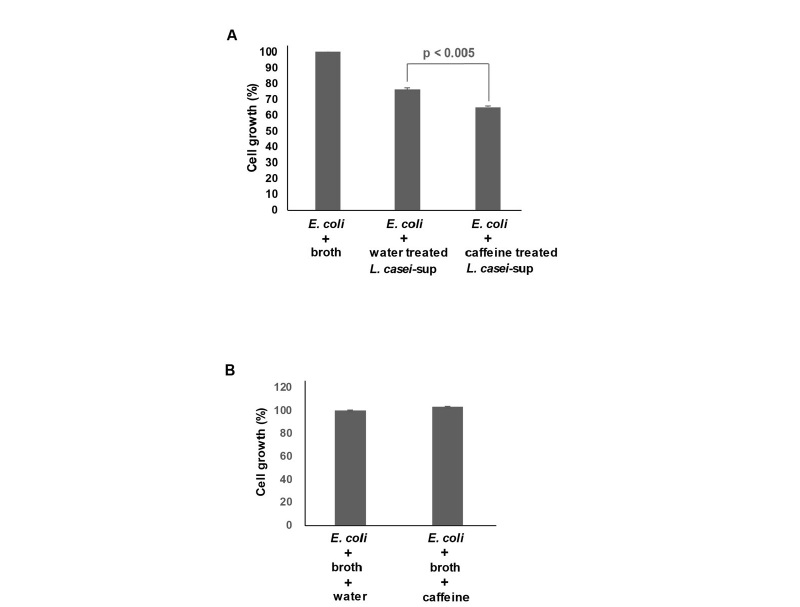
Fig. 1 Effect of caffeine on the antimicrobial activity of L. casei against E. coli. (A) L. casei (3 × 106 CFU/mL) was cultivated in MRS broth in the presence or absence of caffeine (0.3 mg/mL). After 24 h, cell-free culture supernatants were obtained by centrifugation followed by fltration. E. coli (1 × 106 CFU/mL) was inoculated in LB broth together with MRS alone (broth), 10% of the culture supernatant of L. casei (water treated L. casei-sup), or 10% the culture supernatant of caffeine-treated L. casei, and cultivated for 24 h. To measure antimicrobial activity, the optical densities of culture media were measured at 655 nm. The bars represent the means and SEM of the results of three independent experiments, each with triplicate determinations. (B) E. coli (1 × 106 CFU/mL) was inoculated in LB broth together with MRS alone (broth + water) or MRS containing 0.3 mg/mL of caffeine (broth + caffeine). After 24 h, the optical densities of culture media were measured at 655 nm.
L. casei의 젖산 생산과정에 대한 카페인의 영향
L. casei가 생산하는 젖산은 소화관 속 pH를 낮추고 이를 통해 병원성 미생물들의 성장을 지연시킨다고 알려져 있다(Tagg and Dierksen, 2003; Mitchell et al., 2009). 실제로 젖산 생산능력이 L. casei가 나타내는 항균력의 주요 요소로 고려된다(Tagg and Dierksen, 2003). 따라서 연구자 등은 카페인 자극 후 나타나는 L. casei의 증가된 항균력이 젖산 생성량 변화와 연관이 있는지를 확인해보고자 하였다. 이를 위해, 카페인(0.3 mg/mL)을 밤새 처치한 L. casei의 배양상등액에서 pH 변화를 측정하였다. Fig. 2에서처럼, L. casei 전용 배양액인 MRS 배양액 자체의 pH는 약 6.2로 나타났다. L. casei 접종 후 밤새 배양한 배양상등액의 pH는 4.2로 크게 낮아졌다. 하지만 카페인 처치가 pH 변화에 미치는 영향이 없었다. 이 결과는 카페인 자극으로 증가하는 L. casei의 항균력이 젖산 생산과 그로 인한 pH 변화와 무관함을 보여준다.
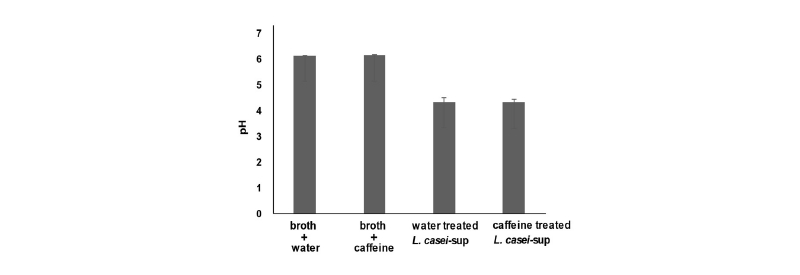
Fig. 2 Role of lactic acid in the ability of caffeine to increase the antimicrobial effect of L. casei. L. casei (3 × 106 CFU/mL) was cultivated in MRS broth in the presence (caffeine treated L. casei-sup) or absence of 0.3 mg/ mL caffeine (water treated L. casei-sup) for 24 h. Cell-free culture supernatants were obtained and the pH of each culture medium was measured. As controls, we also measured the pH of MRS alone (broth + water) and MRS containing 0.3 mg/mL of caffeine (broth + caffeine). The bars represent the means and SEM of the results of three independent experiments, each with triplicate determinations.
카페인 자극 후 L. casei에서 분비되는 항균물질의 열저항성
많은 연구들을 통해서, L. casei의 항균력이 박테리오신 항균펩타이드 분비와 관련이 있다고 알려져 왔다(Ghadbane et al., 2013; Sharma and Srivastava, 2014). 박테리오신은 단백질로부터 생성되고 펩타이드로 분해되어 활성화된다고 알려져 있으며 열저항성이 높은 물질로 알려져 있다(Ghadbane et al., 2013). 이를 토대로, 연구자 등은 카페인 자극 후 L. casei에서 분비되는 항균물질이 박테리오신처럼 열에 대해 높은 저항성을 나타내는지 확인하였다. 이를 위해, 카페인(0.3 mg/mL)을 처치한 후 밤새 배양한 L. casei의 배양상등액을 100℃에서 20분 동안 열처리 하였다. 열처리 상등액을 대장균에 처치하고 대장균의 성장 변화를 확인하였다. 앞서 결과처럼, 카페인 자극 L. casei의 배양상등액은 높은 항균력을 보였다. 하지만 열처리 배양상등액도 동일하게 높은 항균력을 나타내었다(Fig. 3). 이는 L. casei의 항균력을 높여주는 카페인의 효과가 박테리오신과 연관되어 있음을 간접적으로 보여준다.
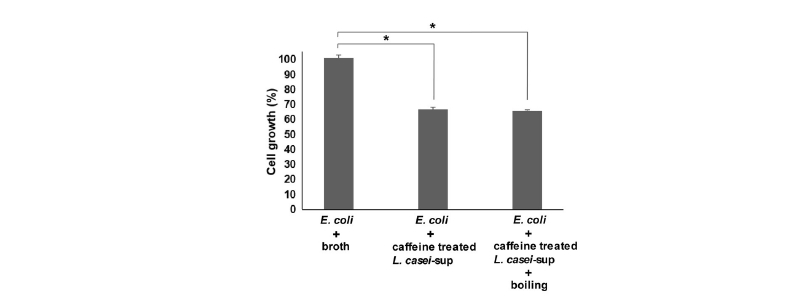
Fig. 3 Nature of an antimicrobial factor present in the culture supernatant of L. casei. E. coli (1 × 106 CFU/mL) was inoculated in LB broth together with MRS alone (broth), the culture supernatant of caffeine-treated L. casei (10%) or the boiled culture supernatant of caffeine-treated L. casei (10%), and cultivated for 24 h. To measure antimicrobial activity, the optical densities of culture media were measured at 655 nm. The bars represent the means and SEM of the results of three independent experiments, each with triplicate determinations (*, p < 0.005).
카페인 자극으로 증가하는 L. casei의 항균 기작
대부분의 항생제들은 세균의 성장과 대사과정에 관련한 효소들을 차단하여 세균을 사멸시키는 것으로 알려져 있다(Falagas et al., 2008). 일부 항생제는 세균의 세포막 형성을 방해하거나(Betrosian and Douzinas, 2009) 직접 세포막에 결합하여 파괴한다(Teng et al., 2014). 연구자 등은 카페인 자극으로 증가하는 L. casei의 항균력이 상대 세균의 세포막 손상과 관련이 있는지 확인해보았다. 이를 위해, L. casei 배양상등액을 대장균에게 처치한 다음 세포막 손상 여부를 PI 염색으로 확인하였다. PI는 손상된 세포막을 통해서만 세포 속으로 유입되는 물질이기 때문이다(Mirnejad et al., 2013). Fig. 4에서처럼, 카페인으로 자극한 L. casei 배양상등액 처치가 대장균의 세포막을 강하게 손상시킴을 확인하였다. 그 손상 정도가 양성대조군으로 사용한 EDTA에 의한 그것과 유사함을 확인하였다. 실제로 여러 종류의 박테리오신들이 상대 균주의 세포막에 소수성 분자로 작용하여 세포막 기능을 저해한다고 알려져 있다(Sharma and Srivastava, 2014). 박테리오신 전사과정이 autoinducer라는 전사 유도성 펩타이드에 의해 조절된다는 사실을 고려하면(Caballero-Guerrero et al., 2013), 카페인이 아마도 autoinducer 펩타이드의 기능 강화를 통해 박테리오신 전사를 증가시키고 이를 통해 유산균의 항균력 증대를 유도했을 것으로 추정된다.
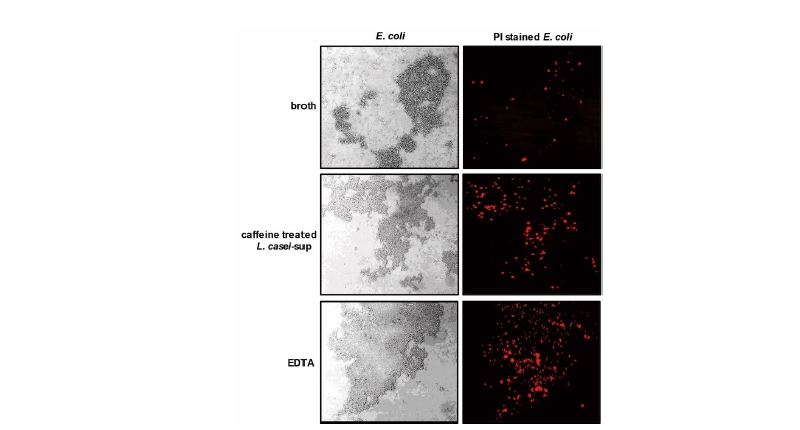
Fig. 4 Culture supernatant of caffeine-treated L. casei causes membrane damage to E. coli. Bacteria (E. coli) were incubated with L. casei culture supernatant for 4 h and then further incubated with PI (100 ng/mL) for 10 min. Bacteria were also incubated with EDTA (the Gram-negative bacteria cell membrane disruptor, 50 mM) as a positive control. Membrane damage was analyzed using a confocal scanning laser microscope (magnification, × 400). The presented results are representative of three independent experiments.
Conclusion
본 연구는 커피의 주성분인 카페인이 장내 유익균으로 알려져 있는 유산균의 항균력에 미치는 영향을 확인하는 것이다. 이를 위해 L. casei에 카페인(0.3 mg/mL)을 처치한 후 배양상등액을 분리하고 항균력을 평가하였다. 그 결과 병원성 대장균에 대한 L. casei 의 항균력이 카페인 처치 후 유의하게 증가함을 확인하였다. 젖산 생산으로 낮은 pH 환경을 유도하는 과정이 L. casei의 주요 항균기작으로 알려져 있지만 카페인 자극으로 증가하는 L. casei 항균력과는 무관하였다. 오히려 L. casei 배양상등액 속 항균물질이 열저항성 특징을 보인다는 사실을 확인하였다. 마지막으로 카페인 자극 L. casei의 배양상등액이 대장균의 세포막을 강하게 손상시킴을 확인하였다. 많은 유산균들의 항균력이 박테리오신이라는 항균펩타이드와 연관이 있다는 점, 그리고 박테리오신이 높은 열저항성과 세포막 손상을 통해 항균력을 나타낸다는 점들을 감안해 볼 때, 본 연구는 L. casei의 항균력을 높여주는 카페인의 효과가 박테리오신의 생산량 증가와 밀접하게 연관되어 있음을 보여준다.
Authors Information
Eunjeong Jang, Division of Life Science and Chemistry, Daejin University, Bachelor
Jin A Park, Division of Life Science and Chemistry, Daejin University, Bachelor
Young Ha Kim, Division of Life Science and Chemistry, Daejin University, Researcher
Ho Kim, Division of Life Science and Chemistry, Daejin University, Professor

