
Introduction
거세는 수소의 고환을 제거하는 방법으로 웅성호르몬의 생성을 현저하게 감소시킨다(Cohen et al., 1990). 웅성호르몬의 감소는 근육내 지방 침착을 향상시켜 육질을 개선하고, 근육의 구조와 질을 변화시켜 연도에 영향을 미친다. 또한 수소의 공격적인 행동을 감소시켜 사양관리를 용이하게 하는 장점이 있다. 한우의 경우 수소의 거세율이 97% 이상으로 나타나고 있으며, 육질 1등급 이상 출현율은 거세우가 88.7%로 수소의 3.5%에 비해 현저히 높은 것으로 보고되고 있다(KAPE, 2020). 이와 같이 거세는 고급육을 생산하기 위한 필수적인 방법이며, 거세 효과를 높이기 위해 거세방법과 거세시기 등 관련 연구가 다양하게 수행되었다(Lee, 2002; Lim et al., 2018). 최근까지 수행된 연구결과에서 수소에 비해 거세우가 증체 및 도체중량에서의 불리함은 있지만, 근내지방도 및 육질이 개선되는 효과가 있는 것으로 검증된 바 있다(Kwak et al., 1995; Kim et al., 2011). 반대로 수소는 증체량이 우수하지만(Jung et al., 1996) 공격적인 행동으로 관리가 어렵고, 근내지방 형성이 저하되기(Cosgrove et al., 1996; Knight et al., 1999) 때문에 경제적으로 불리하다.
따라서 본 연구는 완전 및 반거세 한우의 육성·비육기 성장특성, 혈중 대사물질과 호르몬 및 도체특성을 비교하여 효율적인 한우 고기 생산을 위한 기초자료를 제공하기 위해 수행되었다.
Materials and Methods
본 연구는 국립축산과학원 동물실험윤리위원회의 운영규정에 의해 검토 및 승인 받았으며(NIAS-20181355), 시험동물의 관리 및 실험 절차는 동물실험윤리 위원회의 규정을 준수하였다.
공시동물 및 시험설계
공시동물은 생후 6개월령 한우 송아지 12두(평균체중 153.2 ± 21.0 kg)를 이용하였으며, 시험구 처리는 완전거세(complete castration, CC)와 한쪽 고환만 제거하는 반거세(hemi castration, HC) 방법으로 2처리 하였고, 생후 7개월령에 외과적 수술법으로 거세를 실시하였다.
사양관리
시험축은 단기비육 사양관리 프로그램을 적용하여 육성기(생후 6 - 14개월령) 및 비육기(생후 15 - 24개월령)로 사육하였다. 육성기에는 육성기용 배합사료(3 - 7.5 kg) 및 국내산 양질 조사료인 이탈리안라이그라스 건초(3 - 4 kg)를 급여하였으며, 비육기에는 비육기용 배합사료(8 - 9.5 kg) 및 볏짚(1 - 3 kg)을 1일 2회로 나누어 정량 급여하였다. 물과 미네랄 블록은 자유롭게 이용할 수 있도록 하였으며, 비육기부터 요결석 방지를 위해 염화암모늄(20 g/두)을 매월 5일간 연속적으로 급여하였다. 기타 사양관리는 국립축산과학원 한우연구소의 사양관리 매뉴얼을 준수하였다.
시험사료의 화학적 조성
시험사료의 화학적 조성을 분석하기 위해 1 mm의 입자크기로 분쇄하였다. 건물, 조단백질, 조지방, 조섬유 및 조회분은 AOAC (2005) 방법으로 분석하였으며, NDF (neutral detergent fiber) 및 ADF (acid detergent fiber)는 Van Soest et al. (1991) 방법으로 분석하였다. 공시사료의 화학적 조성은 Table 1과 같다.
Table 1. Chemical composition of the experimental diets (as-basis).
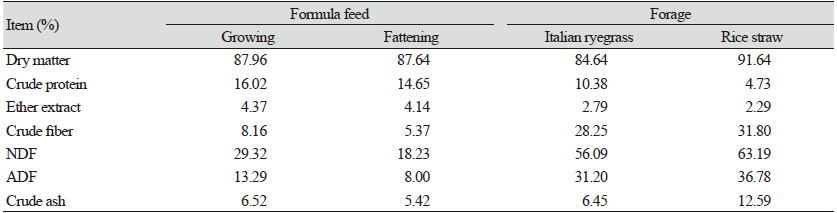
|
|
NDF, neutral detergent fiber; ADF, acid detergent fiber. |
성장특성
시험축의 체중은 시험개시 및 개시 후 매월 1회 오전사료 급여전에 우형기를 이용하여 측정하였으며, 체중과 사양일수를 근거로 일당증체량을 계산하였다. 건물섭취량은 매일 급여된 사료의 잔량을 측정하여 산출하였고, 사료요구율은 일당증체량과 건물섭취량을 이용하여 계산하였다.
혈중 대사물질 및 웅성 호르몬
혈액은 오전 사료 급여전에 시험축의 경정맥으로부터 18 gauge needle 및 진공채혈관(Vacutainer; Becton-Dickinson, NJ, USA)을 이용하여 약 10 mL를 채취하였다. 채취한 혈액은 1,250 × g 에서 20분간 원심 분리하여 혈청을 분리한 후 -80℃에 보관하였다. 이후 분리된 혈청을 이용하여 자동혈액분석기(Hitachi 7020, Hitachi Ltd., Tokyo, Japan)로 혈중 대사물질을 분석하였으며, 분석항목은 glucose (GLU), non-esterified fatty acid (NEFA), albumin (ALB), cholesterol (CHO), triglyceride (TG), blood urea nitrogen (BUN) 이었다.
혈중 웅성호르몬 분석은 Bovine ELISA kit (Diagnostic Products Corporation, Los Angeles, USA)를 이용하여 실시하였다. 호르몬에 대한 특이 항체를 충분히 부착시켜 놓은 plates에 혈청을 첨가하여 반응시킨 다음 I125으로 표지한 이차 항체를 첨가하여 침전시킨 후 흡광도를 측정하여 분석하였다(Gamma-counter, Wallac, UK).
도체특성
도체특성은 사양시험 종료 후 모든 공시축을 축산물공판장에 출하하여 도축 후, 0℃에서 18 - 24시간 동안 도체를 현수 시킨 후 육량 판정요인(도체중, 등지방두께 및 배최장근단면적)과 육질 판정요인(근내지방도, 육색, 지방색, 조직감 및 성숙도)을 축산물 등급판정 세부기준의 소도체 등급판정기준(MAFRA, 2018)에 따라 축산물등급판정사가 판정하였다.
통계분석
본 실험의 모든 결과는 SAS 9.3 program (SAS Institute, Cary, NC, USA)을 이용한 T-test 검정으로 두 집단 간의 평균을 비교하여 5% 수준에서 유의성 검정을 실시하였으며, 통계모형은 다음과 같다.
Yij = µ + treati + eij (1)
여기서, Yij = i번째 고정효과의 j번째 배치, µ = 전체평균, treati = 고정효과(i번째효과: 완전거세, 반거세), eij = 임의오차.
Results
성장특성
완전 및 반거세 한우의 육성·비육기 성장특성은 Table 2와 같다. 개시체중은 처리간 차이가 없었지만, 육성기 종료 체중(14개월령)은 CC에 비해 HC에서 21.2 kg 많았으며, 시험 종료체중(24개월령)도 HC가 CC 보다 66.3 kg 많았다(p < 0.04). 일당증체량은 육성기 및 비육기 모두 CC에 비해 HC에서 높았으며(p < 0.01), 평균적으로 12.6% 증가되었다. 육성기 건물섭취량은 처리간 비슷한 수준이었지만, 비육기에는 HC에서 배합사료 섭취량이 증가되었으며(p < 0.01), 평균 건물섭취량은 CC 보다 HC에서 0.23 kg 많았다. 사료요구율은 CC에 비해 HC에서 육성기 및 비육기 각각 7.7% (p < 0.03) 및 16.8% 낮았으며, 평균 사료요구율도 일당증체량의 영향으로 HC에서 유의적으로 개선되는 결과를 보였다(p < 0.04).
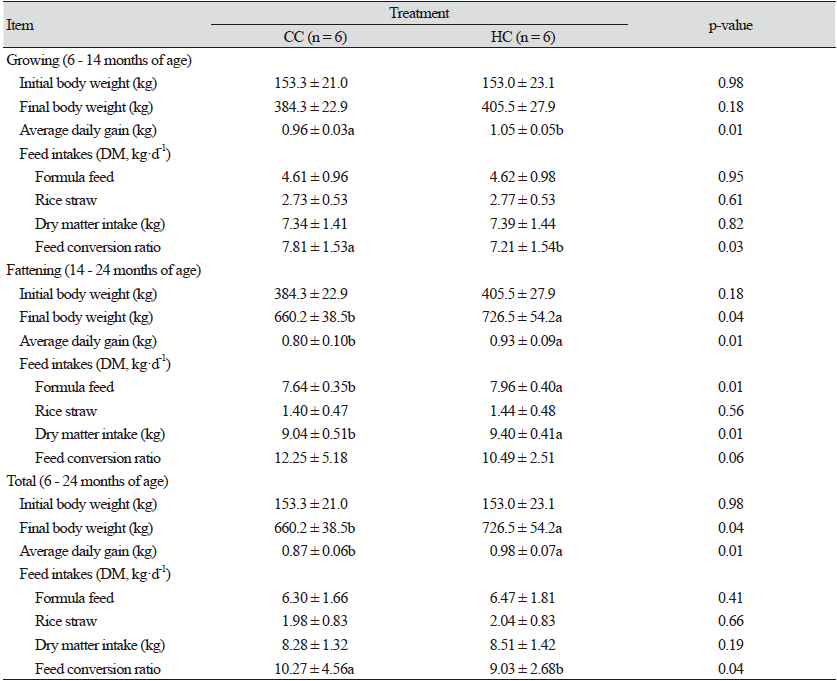
혈중 대사물질
완전 및 반거세 한우의 육성·비육기 혈중 대사물질 농도는 Table 3과 같다. 혈중 GLU 농도는 육성기 동안 처리간 비슷한 수준이었으며, 비육기에는 CC 및 HC 모두 육성기에 비해 농도가 감소되었지만, 처리간 차이는 적었다. 혈중 NEFA 농도는 육성기에는 거세 처리에 관계없이 유사하였고, 비육기에는 HC에 비해 CC에서 감소되는 결과를 보였다(p < 0.01). 육성기 및 비육기 모두 혈중 ALB 농도의 처리간 차이는 없었으며, 혈중 BUN 농도는 CC에 비해 HC에서 낮은 경향을 보였지만 유의적인 차이는 나타나지 않았다. 육성기 혈중 CHO 농도는 HC에 비해 CC에서 유의적으로 높게 나타났으며(p < 0.02), 비육기에도 CC가 HC 보다 높게 유지되었다(p < 0.04). 혈중 TG 농도는 육성기 및 비육기 모두 CC가 HC에 비해 높은 결과를 보였다(p < 0.01).
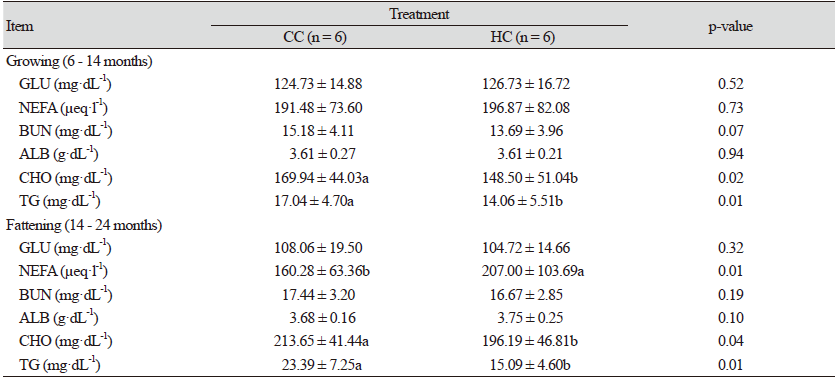
웅성호르몬
완전 및 반거세 한우의 혈중 웅성호르몬(testosterone) 농도 변화는 Fig. 1과 같다. 혈중 웅성호르몬 농도는 거세전에 HC 및 CC간 비슷한 수준이었지만, 거세 후 CC에 비해 HC에서 지속적으로 높게 나타났으며, 9개월령부터 유의적으로 높은 결과를 보였다(p < 0.05). 특히 CC의 경우 거세 후 웅성호르몬 농도가 현저히 감소되어 0.08 - 0.56 ng·mL-1 수준의 농도를 유지하였지만, HC의 경우 소폭 감소된 후 11개월령까지 증가되어 최대 16.51 ng·mL-1의 농도를 보였으며, 이후 증감되는 경향을 보였다.
도체특성
완전 및 반거세 한우의 도체특성은 Table 4와 같다. 도체중은 CC에 비해 HC에서 높았으며, 등심단면적도 넓어지는 경향을 보였지만 통계적인 차이는 없었다. 등지방두께는 HC 보다 CC에서 두껍게 나타났으며(p < 0.02), 육량지수는 CC에 비해 HC에서 유의적으로 높은 결과를 보였다(p < 0.01). 근내지방도는 HC에 비해 CC에서 2배 이상 높게 나타났지만 유의적인 차이는 없었으며, 육색, 지방색, 조직감 및 성숙도는 처리간 비슷한 수준이었다.
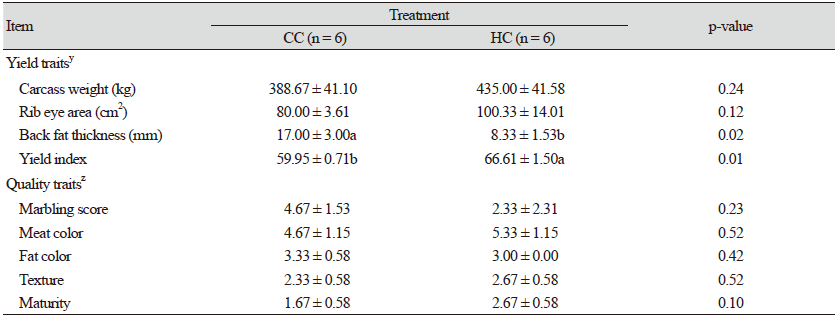
Discussion
한우 수소는 성장과정에서 생식기관의 발달에 따라 성성숙과 관련된 다양한 호르몬들이 분비된다. 그 중에서 웅성호르몬(testosterone)은 성선자극 호르몬인 황체형성호르몬이 고환의 간질세포를 자극함으로써 분비된다(Zirkin and Chen, 2000). 웅성호르몬은 제2차 성징을 유지하고, 질소의 저류를 촉진시켜 단백질 합성을 향상시키며(Gulia et al., 2010), 위성세포 및 근육세포의 증가와 발달을 촉진시키는 것으로 보고되고 있다(Sinha-hikim et al., 2003). 또한, 일부 연구에서 성장호르몬 및 인슐린양성장인자 증가에 영향을 미치는 것으로 알려져 있으며(Parker et al., 1984; Jasper, 1985), Kim et al. (1992) 연구에서도 생식선을 제거한 흰쥐에서 testosterone 투여량과 비례하여 성장호르몬 분비량이 증가되는 것으로 보고된 바 있다. 이와 같이 웅성호르몬은 체성장에 중요한 요인이며, Sundby et al. (1981)은 혈청 웅성호르몬 농도가 높을수록 일당증체량과 체중도 증가된다고 하였고, Jung et al. (1996)는 거세한우에 비해 비거세한우에서 증체율 및 사료섭취량이 각각 19.9% 및 8.0% 증가되었으며, 사료요구율은 10% 감소된 것으로 보고하였다. 그러나 거세를 하게 되면 웅성호르몬 농도가 현저히 감소되고, 단백질 이용효율이 낮아지게 되어 증체가 감소되게 된다(Warwick et al., 1970). 본 연구에서 완전거세의 일당증체량 및 사료요구율이 반거세에 비해 감소된 원인도 거세로 인한 웅성호르몬의 급격한 감소가 원인으로 생각된다. 하지만 반거세의 경우 남아 있는 한쪽 고환에서 웅성호르몬이 분비되어(Fig. 1) 증체와 사료이용효율에 영향을 미친 것으로 판단된다. Barnes et al. (1981)은 수소 및 반거세우간 체중 차이가 없었으며, 반거세우의 경우 한쪽 고환의 크기가 일반적인 수소 고환에 비해 2배 정도 비대해지고, 고환 당 정자수도 증가된다고 하였다. 또한, Sundby et al. (1981)의 연구에서 수소와 반거세우의 성장기 동안 체중 및 웅성호르몬 농도의 차이가 없었으며, Schanbacher et al. (1987)은 수소 및 거세우 보다 반거세우에서 고환의 지름 및 무게가 증가된 것으로 보고하였다. 이러한 연구결과들은 반거세시 남아 있는 한쪽 고환의 발달 및 기능이 향상되어 웅성호르몬이 생리적으로 필요한 수준까지 분비될 수 있음을 시사한다. 본 연구에서도 거세 이후 완전거세구는 웅성호르몬 농도가 감소되어 매우 낮은 수치를 유지하였지만, 반거세구의 경우 일정 수준 이상으로 분비되는 결과를 보여 이전 연구결과와 일치하는 것으로 판단된다. 또한, 10개월령부터 웅성호르몬의 급격한 증가는 성성숙과 관련이 있을 것으로 판단되며, NLRC (2003)는 춘기발동이 시작되는 생후 8개월령부터 성성숙이 최성기에 달하는 12개월령 전후 거세가 가장 경제적인 것으로 보고한 바 있다.
콜레스테롤은 세포막 합성과 유지에 필수적인 물질이며, 특히 웅성호르몬과 같은 스테로이드 호르몬 합성을 위해 필요한 전구물질이다. 콜레스테롤은 P450scc 효소에 의해 21개의 탄소를 가진 pregnenolone으로 전환되고, pregnenolone은 다양한 스테로이드 호르몬을 합성하기 위한 전구체로 사용된다. 콜레스테롤은 사료를 통해 섭취되거나 간에서 중성지방을 이용해 합성되기도 하며, 일반적으로 사료섭취량(배합사료)이 증가되면 혈중 콜레스테롤과 중성지방 농도가 증가되는 것으로 보고되고 있다(Kim, 2018). 본 연구에서도 완전거세 및 반거세 모두 육성기에 비해 배합사료 섭취량이 많은 비육기에 혈중 콜레스테롤과 중성지방 농도가 증가되어 이전 연구결과와 일치하는 것으로 판단된다. 또한, 완전거세에 비해 반거세에서 혈중 콜레스테롤 농도가 감소된 것은 웅성호르몬 합성을 위한 콜레스테롤 이용이 영향을 미친 것으로 사료된다. Lee et al. (2016)은 생후 12, 18, 24 및 32개월령 동안 거세우에 비해 비거세우에서 혈중 콜레스테롤 및 중성지방 농도가 감소되었다고 하여 본 연구와 유사한 결과를 보고한 바 있다.

지방조직의 증가는 지방축적을 통해 지방세포의 크기가 비대해지는 과정과 지방 전구세포의 증식과 분화로 성숙한 지방세포의 수가 증가되는 과정을 통해 일어난다. 한우의 비육 과정에서 피하지방(체지방) 및 근내지방 형성은 모두 성숙한 지방조직의 증가를 통해 이루어지며, 섭취하는 에너지의 양과 월령이 증가될수록 지방조직의 축적이 높아진다. KAPE (2020)의 한우 출하월령별 등급판정 결과에서도 20개월령 이하부터 30개월령까지 육질 1+등급 이상 출현율, 근내지방도 및 등지방두께는 출하월령이 높아질수록 증가되는 것으로 나타나고 있다. 그러나 수소의 경우 출하월령에 따른 차이가 적었으며, 증가율도 낮았다. 또한, Kim and Kim (2017) 연구에서 등지방두께 및 근내지방도는 거세우에 비해 수소에서 현저히 낮게 나타났다. 이와 같은 결과는 웅성호르몬의 지방대사 억제 작용으로 추측되는데, Jeong (2016)은 웅성호르몬이 지방세포에서 중성지방의 축적을 감소시켰으며, peroxisome proliferator-activated receptor γ (PPARγ)와 관련 유전자의 mRNA 발현을 감소시키고, PPARγ의 활성을 억제시킨 것으로 보고하였다. Oh et al. (2005) 연구에서도 testosterone 첨가가 소 근내지방 세포에서 Glycerol-3-Phosphate dehydrogerase (GPDH) 활성을 감소시켜 중성지방 합성을 저해한다고 하였으며, Singh et al. (2003) 연구에서는 testosterone의 첨가 수준이 많아질수록 단위면적당 지방 세포수가 감소되는 것으로 나타났다. 이 같은 결과는 웅성호르몬이 직·간접적인 조절 기전을 통해 지방대사 및 세포를 억제한다는 것을 시사하고 있다. 본 연구에서도 완전거세에 비해 반거세에서 근내지방도 및 등지방두께가 현저히 감소되는 것으로 나타나 한쪽 고환에서 분비되는 웅성호르몬이 지방세포 형성에 부정적인 영향을 미친 것으로 판단된다. 그럼에도 불구하고 KAPE (2020)의 통계연보에서 24개월령 비거세한우의 근내지방도 및 등지방두께는 각각 1.4 및 5.8 mm로 나타나 반거세우의 경우 수소에 비해 육질이 개선될 수 있을 것으로 기대된다.
Conclusion
반거세는 완전거세에 비해 한쪽 고환에서 분비되는 웅성호르몬의 영향으로 증체 및 사료 이용효율이 개선될 수 있지만, 근내지방도가 높은 고급육 생산에는 한계점이 있을 것으로 판단되며, 향후 도체품질 분석을 통해 활용방안에 대한 다양한 검토가 필요할 것으로 판단된다.
Acknowledgements
본 연구는 농촌진흥청 국립축산과학원의 한우 거세방법별 단기비육 기술개발 (PJ01422901) 연구 및 전문연구원 과정 지원사업에 의해 수행되었습니다.
Authors Information
Jun Sang Ahn, https://orcid.org/0000-0001-7362-9270
Sun Sick Jang, https://orcid.org/0000-0002-8121-4697
Ui Hyung Kim, https://orcid.org/0000-0002-2197-5080
So Mi Hwang, https://orcid.org/0000-0002-4152-5984
Jeong Il Won, https://orcid.org/0000-0003-3151-7144
Shil Jin, https://orcid.org/0000-0003-1120-3631
Byung Ki Park, https://orcid.org/0000-0001-5469-2012
Eung Gi Kwon, https://orcid.org/0000-0002-5585-5909

