
Introduction
Ruminants have lower nitrogen efficiency than monogastric species, as dietary crude protein (CP) consumed by them is converted into ammonia by rumen microorganisms and is absorbed in the portal blood (Bach et al., 2005). Only 25% of the nitrogen derived from dietary CP is utilized by the host (Spek et al., 2013), whereas ammonia is changed to urea in the liver and excreted in urine and feces, causing nitrogen waste and adverse environmental effects (Chase et al., 2012).
For decades, scientists made various efforts to improve the nitrogen utilization efficiency, including studies to decrease dietary CP (Frank and Swensson, 2002; Colmenero and Broderick, 2006), feeding protected protein, and feeding protected amino acid (AA) that escape from the degradation of microorganism (Nursoy et al., 2018; Liang et al., 2019). A decreasing dietary CP increases the nitrogen efficiency of ruminants; however, it may compromise animal production (Colmenero and Broderick, 2006; Barros et al., 2017). Although feeding protected protein or increasing rumen-undegradable protein (RUP) meet specific criteria of CP content, it may limit the essential AAs required in the small intestine. Such a phenomenon occurs because AAs composition in the small intestine is largely different depending on the types of protein or the microbial degradation of dietary protein used to synthesize microbial protein (Santos et al., 1998). Recently, studies reported that the protection of the limited AAs improves nitrogen efficiency (Nursoy et al., 2018; Liang et al., 2019).
Several types of rumen-protected AAs (RPAAs) have been developed to improve nitrogen efficiency by resisting the degradation of microorganisms in the rumen, categorized as physical or chemical methods. Physical rumen protection methods include lipid encapsulation (Batistel et al., 2017; Ahn et al., 2019) and pH-sensitive polymers that protect their core at a pH of 5.5 to 7.0 in the rumen and release AAs at a pH of 2 to 3 in the abomasum (Schwab, 1995). These physical methods are characterized by resisting rumen microbial digestive enzymes with low rumen solubility and without AAs interaction (Wu and Papas, 1997). The chemical method includes analog and derivatives of AAs, where the α-amino group is replaced with a non-nitrogenous group or is modified with a chemical blocking to minimize the access by microorganism or enzyme (Schwab, 1995). Recently, interest in RPAA have been increased as some domestic companies are developing RPAA. Most RPAA products are developed through physical methods, however, attempts to protect amino acids using the chemical method are rare. In this study, several AAs were protected based on the chemical methods that were derivatives from a Korean company (CJ CheilJedang, Seoul, Korea). Therefore, the objective of this study was to evaluate the effects of newly developed RPAA prototypes created using chemical methods on rumen fermentation characteristics and examine their protection rate in vitro.
Materials and Methods
All experimental procedures were approved and performed under the guidelines of the Institutional Animal Care and Use Committee of the Kyungpook National University (Approved No. 2020-0129).
Experimental diets and treatments
In this study, timothy hay and a commercial concentrate ground to 1 mm were used as substrate diets. The RPAA prototypes consisted of methionine (RPMet), tryptophan (RPTrp), valine (RPVal), phenylalanine (RPPhe), leucine (RPLeu), and histidine (RPHis) obtained from CJ CheilJedang, Korea. Each RPAA prototype has a similar chemical structure, acetylated AA, with an acetyl group chemically conjugated to the amine group (NH2-) of relevant AA. This acetyl moiety acts as a protection group that can inhibit AA utilization by rumen microbes or enzymes. Proximate analysis of substrate feed and RPAAs are shown in Table 1. The AA compositions of the substrate feed are shown in Table 2.
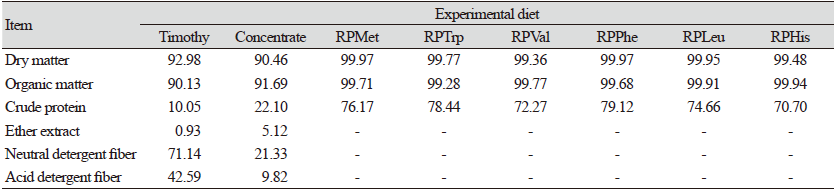
Table 2. Amino acid composition of experimental diets (mg·g-1).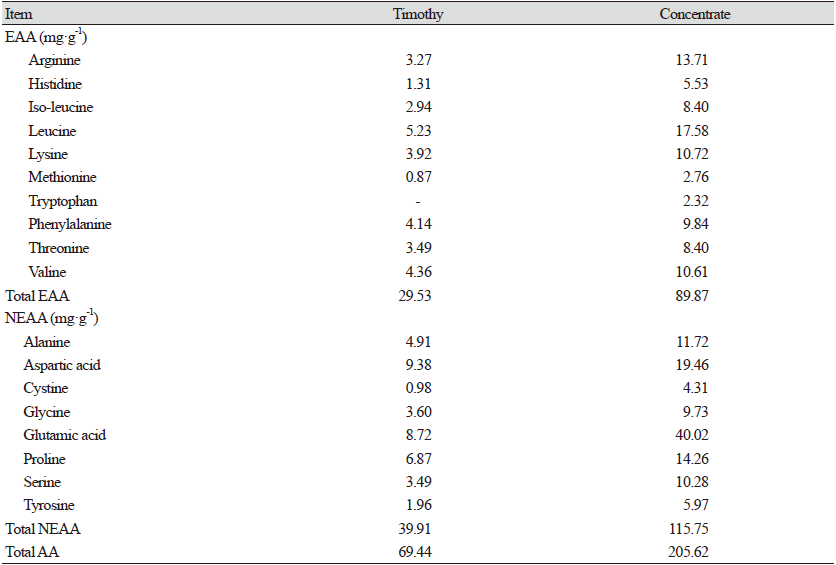
|
|
EAA, essential amino acid; NEAA, non-essential amino acid;A A, amino acid. |
Measures of 0.24 g of timothy hay and 0.16 g (as-fed basis) of a commercial dairy cow concentrate (FARMSCO Inc., Anseong, Korea) were placed in each serum bottle. This ratio is referred to as the ratio used in typical Korean dairy farms. Nine experimental conditions were designed to have different RPAA prototypes and the level of supplementation: 1) control (CON; no RPAA prototype supplement), 2) 0.5% RPMet, 3) 0.5% RPTrp, 4) 0.5% RPVal, 5) 0.5% RPPhe, 6) 0.5% RPLeu, 7) 0.5% RPHis, 8) 20% RPMet, and 9) 20% RPTrp. A high supplementation level (i.e., 20% of the substrate used) was set as an experimental treatment to examine the obvious effect of the specific RPAAs.
In vitro rumen simulation study
Rumen fluid was collected from two Hanwoo cows using a stomach tube at 07:00 h, before the morning feeding (07:30 h) at the experimental farm of the university. The rumen fluid was pooled and immediately transported to a laboratory. Then, it was filtered through eight layers of muslin purged with CO2 to maintain anaerobic conditions. The filtered rumen fluid was mixed with McDougall’s buffer (McDougall, 1948) at a ratio of 1 : 4. Then, 50 mL of the rumen inoculum was filled into each serum bottle containing substrate diets. Later, each serum bottle was capped using a rubber stopper and an aluminum cap. The serum bottles were incubated in triplicates for 0, 6, and 24 h at 39℃.
Analysis of experimental diets and rumen fermentation
Samples were analyzed for dry matter (method 934.01), organic matter (method 942.05), ether extract (method 920.39A), CP (method 2001.11), neutral detergent fiber (method 2002.04) with alpha-amylase and sodium sulfite, and acid detergent fiber (method 973.18), according to AOAC (AOAC, 2019). At the end of each incubation, the serum bottles were opened and poured into a 5 × 10 cm nylon bag (pore size 50 µm, R510, ANKOM Technology, USA), and the pH of the rumen fluid filtered from the nylon bag was analyzed using a pH meter (ST210, Ohaus, New Jersey, USA).
Residues that remained in the serum bottles were poured into nylon bags to determine their dry matter digestibility (DMD). The nylon bag was rinsed with distilled water until the water became clear and dried overnight at 105℃. The ammonia-N was analyzed according to the method described in Chaney and Marbach (1962). A total of 20 µL of sample was mixed with 1 mL of phenol color regent (phenol 50 g and sodium nitroferricyanide 0.25 g per 1 L) and 1 mL of alkali hypochlorite reagent (sodium hydroxide 25 g and sodium hypochlorite 16.8 mL per 1 L). Then, the mixture reacted for 30 min at room temperature. The ammonia-N concentration was analyzed colorimetrically using a spectrophotometer (Optizen pop, Mecasys, Daejeon, Korea) at 630 nm wavelength.
Volatile fatty acids (VFAs) were analyzed with the method by Erwin et al. (1961) using gas chromatography (450-GC, Bruker Inc., Billerica, USA) equipped with BR-Wax fame (BR87503, Bruker Inc., Billerica, USA). Injector, detector (flame ionization detector), and oven temperature were 250, 250, and 100℃, respectively. The airflow for nitrogen, hydrogen, and air were set at 29, 30, and 300 mL·min-1, respectively.
Calculation of rumen protection rate
The rumen protection rate of the RPAA prototypes was calculated based on the relative concentration ratio (%) between 0 h and 6 h or 24 h. The RPAA concentration was analyzed at the CJ CheilJedang company. Briefly, the incubated rumen fluid was centrifuged (1730MR, Labogene, Seoul, Korea) for 10 min at 9,425 × g, and the supernatant was separated and filtered using a 0.45 µm syringe filter (RJN1345NH, Rephile, Shanghai, China). Then, the RPAA prototype concentration in rumen fluid was analyzed with an HPLC (CBM-20A, Shimadzu, Kyoto, Japan) equipped with a Purospher® STAR RP-18 end-capped 5 µm column (EMD Millipore, Darmstadt, Germany) at 230 nm wavelength. Distilled water, 0.2% H3PO4, C2H3N, and 0.2% H3PO4 were used as the pump solvents.
Statistical analysis
All data were subjected to analysis of variance, and further mean comparisons were performed with Duncan’s multiple range test to determine significant differences using SPSS (IBM Corporation, 2017). The treatment effect was analyzed; however, the incubation time effect was not statistically examined. Differences were considered significant at p < 0.05, and p-values between 0.05 and 0.10 were considered to represent a tendency.
Results and Discussion
The effects of the RPAA prototypes developed by a chemical method were evaluated on in vitro rumen fermentation and protection rate in this study. Due to the lack of derivatives in vitro rumen fermentation research, we referred to 2-hydroxy-4-(methylthio) butanoic acid (HMB) and isopropyl ester form of HMB (iHMB), which was one of the chemical rumen-protected methods. Briefly, 50% of these are bypassed in the rumen and absorbed in the small intestine, converted into keto methylthio butyrate that is intermediate to synthesize Met and transaminated to L-Met that the host animal can utilize for maintenance, growth, and production (Firkins et al., 2015).
The rumen fermentation in vitro results are shown in Tables 3 and 4. The pH was lower in the CON (6.88 and 6.79 at 6 and 24 h, respectively) than the other treatments at 6 h and 24 h (p < 0.05; Table 3). Dry matter digestibility was higher in the 20% RPMet and the 20% RPTrp treatments than the other groups at 6 h and 24 h (p < 0.05). However, except for the 20% RPMet and the 20% RPTrp treatments, there was no significant difference among groups in DMD (Table 3). The DMD is used as a fermentation indicator in feed evaluation when using in vitro rumen simulation. At the end of each fermentation process, filter crucible (Aderao et al., 2018), filter paper (Navarro-Villa et al., 2011), or Ankom filter bag (Hart et al., 2009) were often used to filter the substrate residues and further to dry it in an oven to determine the dry matter of the residues in the serum bottle. In our study, the DMD was determined using nylon bags (Guo et al., 2019). The DMD of all treatments was not significant, except for the RPMet 20% and the RPTrp 20% treatments (Table 3). The DMD results were similar to those observed in previous research (Vazquez-Anon et al., 2001), with the addition of the HMB concentration (0, 0.20, 0.77, and 1.43% in the diet) in in vitro rumen fermentation, except for the RPMet 20% and the RPTrp 20% treatments. Nevertheless, both the RPMet 20% and the RPTrp 20% treatments had higher DMDs than the rest of the groups (p < 0.05). The reason behind this result may be that the nylon bag used for determining the DMD had a 50 µm pore size, and the RPAAs used here were high solubility in water and had a small particle size, such as fine granules. In the in vitro rumen fermentation, the residue in nylon bags is considered as indigestion. Thus, the DMD was calculated by subtracting the residue (%) from the total percentage (100%). Probably, when the rumen fluid was distributed in bottles of the RPMet 20% and the RPTrp 20% treatments, their high concentration level occurred because the RPAA prototypes were either soluble in the rumen fluid or passed through the nylon bag during the filtering process. Therefore, the DMD was likely to be overestimated (Table 3). Our results indicate that using nylon bags or any other filtering methodology during in vitro rumen simulation may not be an appropriate method to estimate the effect of RPAA on DMD, especially when the solubility or particle size of the RPAA is being evaluated. In line with the above-mentioned hypothesis, Bach et al. (2005) also noted that high solubility in rumen fluid did not result in high rumen degradability, and the rumen degradability degree depended on molecular structure.
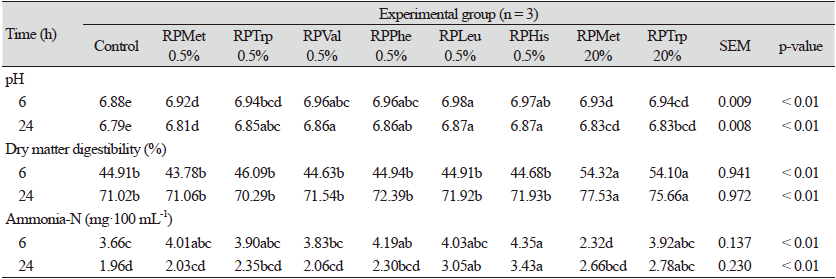
The ammonia-N of the CON treatment showed the lowest level among the groups (p < 0.05), except for 20% RPMet at 6 h (p < 0.05). The protection rate of 20% RPMet at 6 h showed higher rate (93.75%). The lowest ammonia-N concentration in 20% RPMet at 6 h was one of the peculiar phenomena in this study. It was expected that with high protection rate of RPMet, the ammonia-N concentration of the supernatant would be similar or a little higher compared with the control and yet the results were somewhat different. It is not clear why such results were obtained. Further experiments are planned to elucidate this phenomenon.
The total VFA concentration of the 20% RPMet (27.95 and 52.56 mmol at 6 h and 24 h, respectively) was higher than the rest of the groups at 6 and 24 h (p <0.05). The 20% RPMet supplementation had the highest molar proportion of acetate but the lowest proportion of propionate compared to other groups at 6 h and 24 h (p < 0.05). Consequently, acetate : propionate (A : P) ratio was highest in the 20% RPMet (Table 4). Volatile fatty acid, one of the in vitro rumen fermentation indicators, supplies more than 70% of the energy to ruminants (Bergman, 1990). The acetate level, which accounts for the largest percentage of VFA, was relatively higher in the 20% RPMet and the 20% RPTrp treatments. In the case of branched-chain VFA (BCVFA), it is produced when protein and AAs, especially branched-chain AAs (BCAAs), are degraded (Allison and Bryant, 1963). Therefore, the BCVFA concentration has been used as another indicator in RPAA-related studies (Fowler et al., 2015; Baghbanzadeh-Nobari et al., 2017). Previous research showed that BCVFA promotes the growth of cellulolytic microbes in the rumen (Dehority et al., 1967) and increases total VFA and acetate concentrations (Liu et al., 2009). However, in this study, although the concentration of BCVFAs in the 20% RPMet treatment was not higher than other treatments, and acetate was the highest in this treatment (p < 0.05). The results of studies to evaluate the effects of RPAA on VFA are limited and controversial. The study by Vazquez-Anon et al. (2001), who supplemented 0, 0.2, 0.77, and 1.43% HMB in in vitro rumen fermentation, did not find significant differences in the total VFA and individual VFA. In agreement with this study, Baghbanzadeh-Nobari et al. (2017), who supplemented 1.8 g·kg-1 HMBi to ewe, found an increase in the total VFA, acetate, and A : P ratio. In addition, Noftsger et al. (2003), who supplemented 0, 0.055, and 0.11% HMB in in vitro rumen fermentation, found a significant propionate decrease. Notably, it was confirmed that the propionate concentration decreased as the RPAA concentrations increased. The effect of RPAAs on VFA concentration was not apparent.
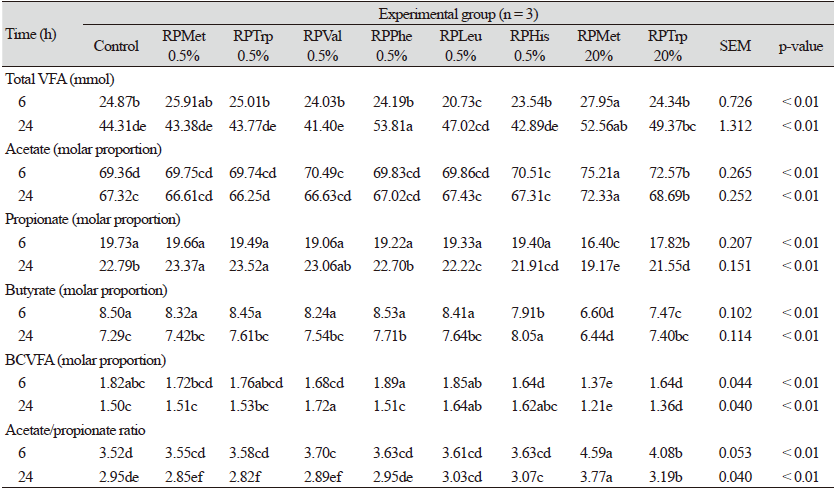
Table 5 shows the in vitro rumen respective AA contents in the different incubation hours and the rumen and protection rate of the AAs at 6 h and 24 h. The in vitro rumen protection rate of the 0.5% RPMet and the 0.5% RPTrp treatments were 29.85% and 40.28%, whereas the 20% RPMet and the 20% RPTrp treatments showed 90.70% and 109.21% protection rates at 24 h, respectively. Other prototypes showed 75.05% (RPVal), 63.77% (RPPhe), 73.41% (RPLeu), and 78.74% (RPHis) of protection rates at 24 h. The protection rates of the RPAAs prototypes were not significantly different among groups at 6 h, except for the 0.5% RPMet and the 20% RPTrp treatments, and the 20% RPTrp showed the highest protection rates among the RPAA prototypes at 24 h.
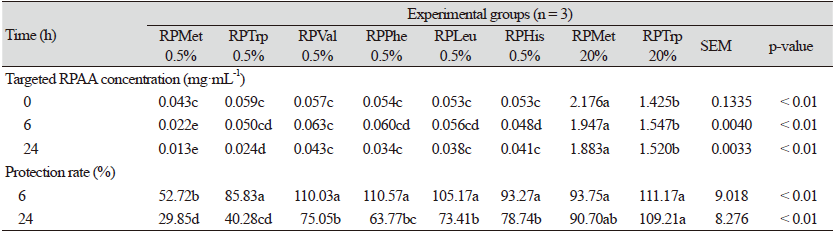
The methods used for evaluating the protection rate of RPAAs were different depending on the protection methods. In the case of coating-based RPAA, for example, in situ experiments (Berthiaume et al., 2000; Koenig and Rode, 2001), which uses nylon bags, are regarded as a standard approach. However, the RPAA prototypes used here have some unique characteristics compared to conventional coating-based RPAAs, such as 1) powder form, 2) relatively soluble in rumen fluid, and 3) split of prototypes in NH2-blocking group and AA in the small intestine by enzyme (Endo, 1980; Windschitl and Stern, 1988). Using nylon bags via an in situ methodology to estimate the protection rate of RPAA developed by the chemical method is likely to underestimate the actual protection rate. Because the RPAA solubility was different (data not shown), it is improbable that the protection rate of RPAA prototypes was analyzed using residue in nylon bag (Wilman and Adesogan, 2000; Schwab and Ordway, 2003). Therefore, our study was conducted using batch culture in vitro to evaluate the rumen protection rate. in vitro rumen protection rates of the 0.5% RPMet and the 0.5% RPTrp treatments were 29.85% and 40.28%, whereas the 20% RPMet and the 20% RPTrp treatments were 90.70% and 109.21% at 24 h, respectively. Especially, the protection rate of RPMet and RPTrp was affected by the level of supplementation (0.5% or 20%). It is not clear why the protection rate was different depending on its supplementation level. Velle et al. (1997) reported that rumen degradation was reduced as methionine dosage increased (75, 150, 300, and 600 mmol), although unprotected methionine was used in the study. They further discussed that such discrepancy has occurred because the liberation of sulfur inhibits deaminating enzymes. Another study has also reported that derivatives were not adapted to rumen microbes as AAs (Witt et al., 1998). Thus, it may be feasible to conclude that a high derivatives level caused rumen microbes to fail to degrade, resulting in high protection rate. In the case of RPAA products on the market, they have a different level of rumen protection: 90% of coating RPAA with pH-sensitive polymer (Robert and Williams, 1997), 50% of HMBi, another type of AA analog (St-Pierre and Sylvester, 2005), and the 66% or 78% of coating RPAA based on ethyl-cellulose (Berthiaume et al., 2001; Koenig and Rode, 2001). Therefore, the results of this study showed a reasonable range of rumen protection compared to other RPAAs in the literature with 75.05% for RPVal, 63.77% for RPPhe, 73.41% for RPLeu, and 78.74% for RPHis at 24 h.
Conclusion
In this study, the effects of RPAA prototypes, which were chemically synthesized, were evaluated on in vitro rumen fermentation and protection rate. Differences among treatments in terms of ammonia-N and VFA were observed. No DMD differences were noted among treatments, except for the RPMet 20% and the RPTrp 20% supplementations. The in vitro ruminal protection rate of 0.5% of PRMet, PRTrp, PRVal, PRPhe, PRLeu, PRHis, and 20% of RPMet, RPTrp prototypes averaged 29.85, 40.28, 75.05, 63.77, 73.41, 78.74, 90.70, and 109.21% respectively. Especially, the ruminal protection rate of Met and Trp was affected by the level of supplementation (0.5% and 20%), although the reason for such discrepancy remains unclear. Given the in vitro protection rate of RPAA prototypes, derivatives developed in this study have potential as rumen-protected amino acids. However, In vivo studies are needed for the accurate evaluation of the RPAA prototypes.
Acknowledgments
This research was supported by Korea Institute of Planning and Evaluation for Technology in Food, Agriculture and Forestry (IPET) through Agri-Bio industry Technology Development-Program, funded by Ministry of Agriculture, Food and Rural Affairs (MAFRA) (117030-3). This research was also supported by Kyungpook National University Development Project Research Fund, 2018.
Authors Information
Gyeongjin Kim, https://orcid.org/0000-0003-2202-126X
Tabita Dameria Marbun, https://orcid.org/0000-0002-3360-9715
Jinhyun Park, https://orcid.org/0000-0001-6328-4125
Sang Moo Lee, https://orcid.org/0000-0003-2510-2591
Hong Gu Lee, https://orcid.org/0000-0002-0679-5663
Jun Ok Moon, https://orcid.org/0000-0002-2118-3833
Jin Seung Park, https://orcid.org/0000-0002-4333-1024
Eun Joong Kim, https://orcid.org/0000-0002-5962-6994

