
Introduction
동북아시아 지역이 기원지인 콩(Glycine max L.)은 세계 3대 재배작물 중 하나이고, 단백질, 지질, 식이섬유와 같은 주요 영양 성분이 풍부하며 이소플라본, 안토시아닌, 사포닌, 올리고당과 같은 기능성 성분들도 포함하고 있어서, 전 세계적으로 소비와 수요가 지속적으로 증가하고 있는 건강한 식품 소재이다(Lee et al., 2011). 또한 제초제저항성 유전자를 콩에 도입함으로써 상업을 목적으로 콩을 경작하는 것이 증가하였고 크게 성공했다(Pratap et al., 2012). ISAAA (international service for the acquisition of agri-biotech applications)의 보고에 의하면 2019년을 기준으로 GM 작물(genetically modified crops)은 전 세계 29개국에서 재배 면적이 1억 9,040만 헥타르에 이르렀으며, 특히 유전자변형(genetically modified, GM) 콩은 그 중 약 50%인 9,190만 헥타르의 면적에서 재배되고 있다(ISAAA, 2020).
GM 작물은 종자 자체로도 경제적 가치가 있지만, 사회적, 환경적 측면에서도 경제적인 효과가 높다. PG Economics의 보고에 따르면 1996년부터 2016년까지 GM 작물 재배가 시작된 이후 21년간의 세계 농업을 분석한 결과 GM 작물을 재배함으로써 농약사용량, 노동력 절감 및 생산성 증대 등으로 농가소득이 증가하였으며, 탄소배출 감소 및 온실가스 감소의 효과와 생물다양성 보전 등으로 환경을 보호하는 데 효과를 높일 수 있는 등 긍정적인 보고가 있었다(Brookes and Barfoot, 2018). 또한 아직까지 GM 작물 재배로 인해 일반적인 환경에 위협적인 경우가 없었으며, 오히려 기존의 농업 시스템인 일반적인 농작물에 다량의 화학물질을 사용하는 것에 의한 환경 피해가 더 크다고 보고된 바 있다(Raven, 2010; Romeis et al., 2013). 현재 국내에서는 비타민E 강화콩, 화장품소재용 TRX (thioredoxin), EGF (epidermal growth factor), IGF (insulin‐like growth factor) 생산 GM 콩 등의 다양한 기능성 GM 콩이 개발되었다는 보고가 있으며, GM 작물에 대한 연구개발이 지속적으로 수행되고 있으나, 아직까지 상업적인 목적으로 재배되는 GM 작물은 없는 실정이다(Amin et al., 2020a; Kim et al., 2020a; Oh et al., 2020a). 이러한 GM 작물의 긍정적 측면과 지속적인 재배의 증가에도 GM 작물의 잡초화, 잠재적 독성 및 새로운 종류의 독성물질 생성 등으로 인한 생태계 교란 및 환경에 미치는 영향 등 GM 작물의 연구개발에 대해 많은 우려가 계속해서 나오고 있다(Oh et al., 2020b).
GM 작물의 안전성 평가에서 경작지, 주변 환경 및 생태계에 미치는 영향 평가가 필수 항목으로 요구되고 있으며, GM 작물의 개발과 재배가 증가함에 따른 경작지와 주변의 생태학적 문제 예방 및 해결을 위한 다양한 연구들이 수행되었다. 자연 환경 조건에서 GM 콩의 교잡률이 낮더라도 GM 콩에서 야생종으로의 유전자 이동에 의한 생태계 교란 및 다양성 감소 등의 부정적인 결과가 발생될 수 있기 때문에 GM 콩의 환경위해성에 대한 평가의 필요성이 대두되었다(Lu, 2004; Mizuguti et al., 2010). 2017년도에 국내에서 GM 유채와 GM 목화가 비의도적 환경 방출된 사례들이 보고되었고, 핀란드에서는 자연 환경에서 GM 페튜니아가 발견되는 등의 비의도적인 환경 방출 사고들이 발생하고 있다(KBCH, 2018). 이러한 생태계 교란 및 환경에 미치는 영향 등을 분석하기 위해 GM 작물에 대한 환경위해성 평가가 필요하며, 이 평가는 재배지와 그 주변 생태계를 구성하는 생물종을 정량적으로 분석하는 생물다양성(biodiversity) 조사가 선행적으로 수행되어야 한다(Choi et al., 2015). 생물다양성 평가는 단순히 종의 멸종률이나 목록을 다루는 것이 아니라, 인구와 주변 생태계의 변화에 중점을 둔다(Kwon, 2008). 다양한 생물 분류군 중 곤충은 전 세계적으로 백만 종이 넘으며 매우 다양하고 그로 인해 다양한 생태를 가지고 있어 먹이사슬에서 양적으로 우위에 있다(Borror et al., 1989; Groombridge, 1992; Price, 1997). 따라서 곤충은 생태계 먹이연쇄에서 생태계의 특성을 나타내는 데 매우 중요하며, 생태계를 구성하는 곤충의 종 구성변화는 장기적인 생태계의 변화를 파악할 수 있는 중요한 구성요소이다(Gullan and Cranston, 2005). 또한 일반적으로 주변환경과 밀접한 관계를 맺고 해당 생태계 환경의 질을 측정할 수 있는 기준으로 활용할 수 있다(KNPS, 2006).
국내에서 재배 품종인 광안(Glycine max L. cv. Gwangan)콩에 카로티노이드 생합성 유전자인 Psy와 CrtI 유전자를 형질 전환하여 종자에서 카로티노이드 함량이 광안콩보다 약 62배 증가한 비타민A 강화콩이 개발되었다(Kim et al., 2012). 본 연구에서는 GM 콩이 재배 환경에서 거미류를 포함한 곤충상에 미치는 영향을 평가하기 위해, GM 콩인 비타민A 강화콩과 모품종인 광안콩을 LMO (living modified organism) 격리포장에서 재배하고 곤충상의 변화양상을 조사 분석하였으며, 이는 국내에서 연구개발 중인 비타민A 강화콩의 안전성 평가를 위한 기초 자료를 제공할 것이다.
Materials and Methods
재배 정보
콩 재배지의 곤충상 조사를 위해 2019년과 2020년에 비타민A 강화콩과 모품종인 광안콩을 경북대학교 군위 LMO 격리포장(경북 군위군, RDA-가A-2015-049)과 국립농업과학원 전주 LMO 격리포장(전북 전주시, RDA-가AB-2013-041)에 각 품종별로 24 m2 (8 m × 3 m)을 3반복 배치(72 m2)하여 재배하였다. 공시계통인 비타민A 강화콩과 광안콩의 파종과 정식은 2019년에는 각 시험군의 종자 500립씩 50공 트레이에 6월 3일에 파종한 후 6월 25일에 LMO 격리포장에 각 품종별 300주씩 정식하였고, 2020년에는 각 시험군의 종자 500립씩 50공 트레이에 6월 3일에 파종한 후 6월 23일에 LMO 격리포장에 각 품종별 300주씩 정식하였다. 콩의 재배방법은 농촌진흥청의 농사시험 표준재배법을 기준으로 하였다.
채집 시기 및 방법
콩 재배지의 곤충 조사시기는 비타민A 강화콩과 모품종인 광안콩을 대상으로 2019년도 전주 LMO 격리포장에서는 콩 품종의 성장기(8월 9일)부터 수확 직전(10월 4일)까지 2주 간격으로 총 5회에 걸쳐 실시하였으며, 군위 LMO 격리포장에서는 8월 6일부터 10월 1일까지 2주 간격으로 총 5회에 걸쳐 실시하였다. 2020년도 전주 LMO 격리포장에서는 8월 5일부터 9월 29일까지 2주 간격으로 총 5회에 걸쳐 실시하였으며, 군위 LMO 격리포장에서는 8월 5일부터 9월 29일까지 2주 간격으로 총 5회에 걸쳐 실시하였다. 곤충 채집은 각 품종별 3반복으로 수행하였고, 비타민A 강화콩과 모품종인 광안콩 간의 해충, 천적, 일반 곤충으로 분류하여 발생 양상을 조사하였다. 각 조사 구역별 곤충 조사는 실험구내의 황색 끈끈이 트랩(yellow sticky trap, 350 mm × 250 mm, greenagrotech, gyeongsan, Korea)을 이용하여 각 실험구의 1반복당 1개씩을 곤충상 조사 대상인 콩보다 30 cm 높은 위치에 설치하여 2주간 곤충을 채집한 후, 각 채집된 실험구별로 황색 끈끈이 트랩을 지퍼백에 담아 에틸아세테이트(ethyl acetate)를 이용하여 살충하였다.
해충조사는 해충과 천적 및 기타 곤충을 대상으로 기능군별 발생양상을 분석하였다. 채집된 곤충은 기능별로 해충군, 천적군(거미 포함), 기타 곤충군으로 구분하였는데, 해충군은 콩 해충으로 알려진 총채벌레류, 노린재류 및 나방류 등을 분류 선정하였으며 종(species) 수준에서 동정 계수하였다. 또한 포식 및 기생성 곤충과 거미를 포함하여 천적군으로, 해충군과 천적군을 제외한 나머지 곤충들은 기타 곤충군으로 분류 선정하였으며 과(family) 수준에서 동정 계수하였다.
통계 분석
채집된 곤충의 조사 결과에 대한 통계 분석은 SPSS (23.0.0 for windows, Rel.23.0, 2015., SPSS Inc., Chicago, USA)를 사용하여 p <0.05 수준으로 ANOVA와 LSD (least significant difference) 검정으로 평균간의 유의성 여부를 검정하였고, 채집된 곤충의 기능군별로 우점도 지수(dominance index, DI; McNaugton's dominace index) 및 다양도 지수(diversity index, H'; Shannon-Weaver diversity index), 균등도 지수(evenness index, EI; Pielou index), 풍부도 지수(richness index, RI; Margalef species richness index)를 계산하였다(Choi et al., 2015). 또한, 콩의 품종 및 재배 시기에 따른 곤충상 변화를 분석하기 위해 주성분분석(principal component analysis, PCA)과 직교부분최소자승판별분석(orthogonal partial least squares-discriminant analysis, OPLS-DA)을 수행하였으며, 그 결과를 score plot 및 VIP (variable importance in the prediction) 값으로 나타냈다(SIMCA-P version 12.0; Umetrics, Umeå, Sweden).
Results and Discussion
곤충류 발생현황
군위 LMO 격리포장과 전주 LMO 격리포장에서 2019년과 2020년 2년 동안 콩의 성장기에서 종자 수확 직전까지 비타민A 강화콩과 모품종인 광안콩을 재배한 포장에서 연도별로 총 5회에 걸쳐 채집된 곤충을 해충, 천적 및 기타 곤충의 기능군별 발생현황을 비교 분석하였다. 조사기간 중 13목 81과 207,760개체의 곤충류와 거미류가 채집되었으며, 기능군별로는 해충류 7목 27과 43종 142,224개체, 천적류 8목 23과 29,005개체, 기타 곤충류 8목 36과 36,531개체가 채집되었다. 품종별로는 비타민A 강화콩 재배지에서 105,765개체, 광안콩 재배지에서 101,995개체가 채집되었다. 지역별로 채집된 곤충은 군위 81,918개체, 전주 125,842개체로 전주에서 월등히 높은 조사밀도를 보였다(Table 1).
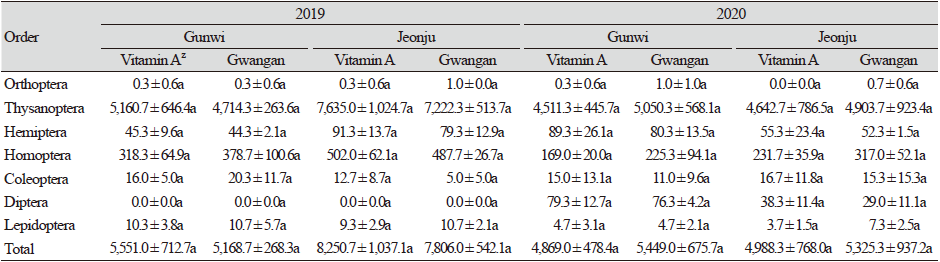
해충류는 군위에서 2019년에 28종 32,159개체, 2020년에 31종 30,954개체가, 전주에서는 2019년에 22종 48,170개체, 2020년에 36종 30,941개체가 조사되었다. 두 지역 모두 2019년보다 2020년에 개체수는 다소 감소하였다. 2019년과 2020년 모두 두 지역에서 꽃노랑총채벌레(Frankliniella occidentalisi)와 대만총채벌레(Frankliniella intonsa)를 포함하는 총채벌레목(Thysanoptera)의 발생밀도가 가장 높았으며 전체 밀도의 대부분을 차지하였다. 2020년 군위에서는 2019년과 달리 대만총채벌레, 애긴노린재(Nysius plebejus), 갈색날개매미충(Ricania shantungensis), 콩잎굴파리(Liriomyza congesta)의 발생밀도가 증가하였으나, 꽃노랑총채벌레, 콩진딧물(Aphis glycines), 막대애매미충(Empoasca rybiogon)은 감소하였다. 2020년에 전주에서는 총채벌레류, 콩진딧물의 발생밀도가 감소하였다(Table 1).
거미류(Araneae)를 포함한 천적류는 2019년 군위에서 15과 6,929개체가, 2020년에는 21과 1,852개체가 조사되었으며, 전주에서는 2019년에 12과 11,748개체가, 2020년에는 21과 8476개체가 조사되어 두 지역 모두 2020년에 발생밀도가 감소하였다. 곤충천적류 중에서는 군위에서 2020년에 머리파리과(Pipunculidae)와 맵시벌과(Ichneumonidae)의 발생이 증가한 반면, 좀벌과(Eulophidae)의 발생은 크게 감소하였으며 꽃노린재과(Anthocoridae), 기생파리과(Tachinidae), 고치벌과(Braconidae)의 발생 역시 감소하였다. 전주에서는 2020년에 꽃노린재과, 풀잠자리과(Chrysopidae), 무당벌레과(Coccinellidae)의 발생이 증가하였고, 반면 좀벌과의 발생이 매우 감소하였으며 기생파리과, 고치벌과의 발생도 다소 감소한 경향을 보였다. 군위에서는 2020년에 거미류 중 꼬마거미과(Theridiidae)가 크게 증가하였으며 갈거미과(Tetragnathidae), 왕거미과(Araneidae)는 감소하였다. 전주에서 깡충거미과(Salticidae)가 증가하였고, 꼬마거미과는 감소하였다. 또한 기타 거미류는 비슷한 발생양상을 나타내었다(Table 1).
해충 및 천적을 제외한 기타 곤충류에서는 2019년에 군위에서 33과 6,393개체가, 2020년에 32과 3,631개체가 조사되어 발생이 감소하였다. 반면, 전주에서는 2019년에 20과 21,540개체가, 2020년에 31과 4,967개체가 조사되어 2019년에 높은 발생밀도를 보였다. 군위에서 2020년 깔따구과(Chiron omidae), 집파리과(Muscidae), 검정파리과(Calliphoridae)의 발생량이 증가하였고, 전주에서 버섯파리과(Mycetophilidae)와 춤파리과(Empididae)의 발생량이 증가하였다. 군위와 전주 모두 2019년에는 나방파리과(Psychodidae)의 발생밀도가 매우 높았으나, 2020년에는 매우 감소하였다. 이외에도 노랑굴파리과(Chloropidae), 매미충과(Cicadellidae), 나무이과(Psyllidae)의 발생도 매우 높았다. 이와 같이 군위와 전주 두 지역 모두 2020년이 2019년보다 천적 및 기타곤충류의 발생량이 감소하였으며, 해충 발생량에서는 전주에서는 감소하였으나 군위에서는 큰 변화가 없었고, 곤충 종류에 따라 발생량에 차이가 나타나기도 하였다(Table 1).
또한, 지역 및 연차간 품종별 비표적 곤충류 발생현황을 분석한 결과, 연도별로는 2019년 군위에서는 광안콩과 비타민A 강화콩에서 각각 22,422개체, 23,059개체가 조사되었으며, 2020년에는 각각 19,218개체, 17,219개체가 조사되어 연차간 큰 차이는 없는 것으로 나왔다. 반면 전주에서는 2019년에 광안콩과 비타민A 강화콩에서 각각 39,704개체, 41,754개체가 발생하였으며, 2020년에는 각각 20,651개체, 23,733개체가 발생하여 2019년에 비해 개체발생량이 현저히 낮아진 것으로 나타났다. 지역별로는 2019년에 전주지역에서 해충을 비롯한 전반적인 개체군밀도가 군위지역보다 더 높게 나타났으나 2020년에는 큰 차이가 나타나지 않았다. 요약하면, 지역간 및 연차간 발생량의 차이가 나타났지만, 품종별로 통계적 유의차는 나타나지 않았다(Table 1; Fig. 1).
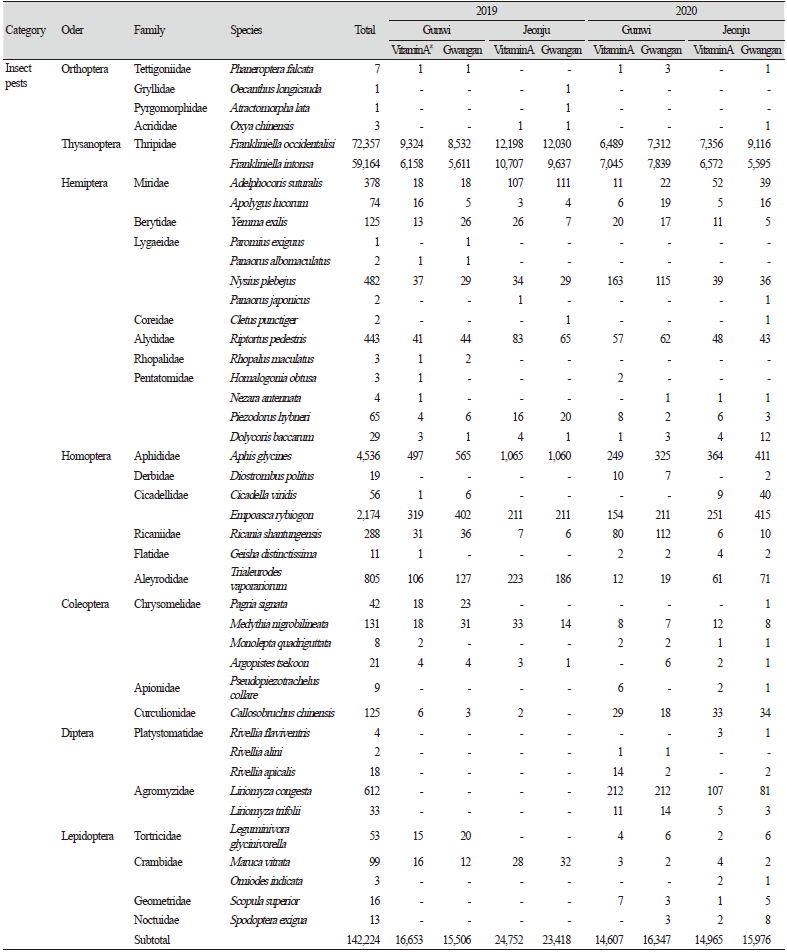
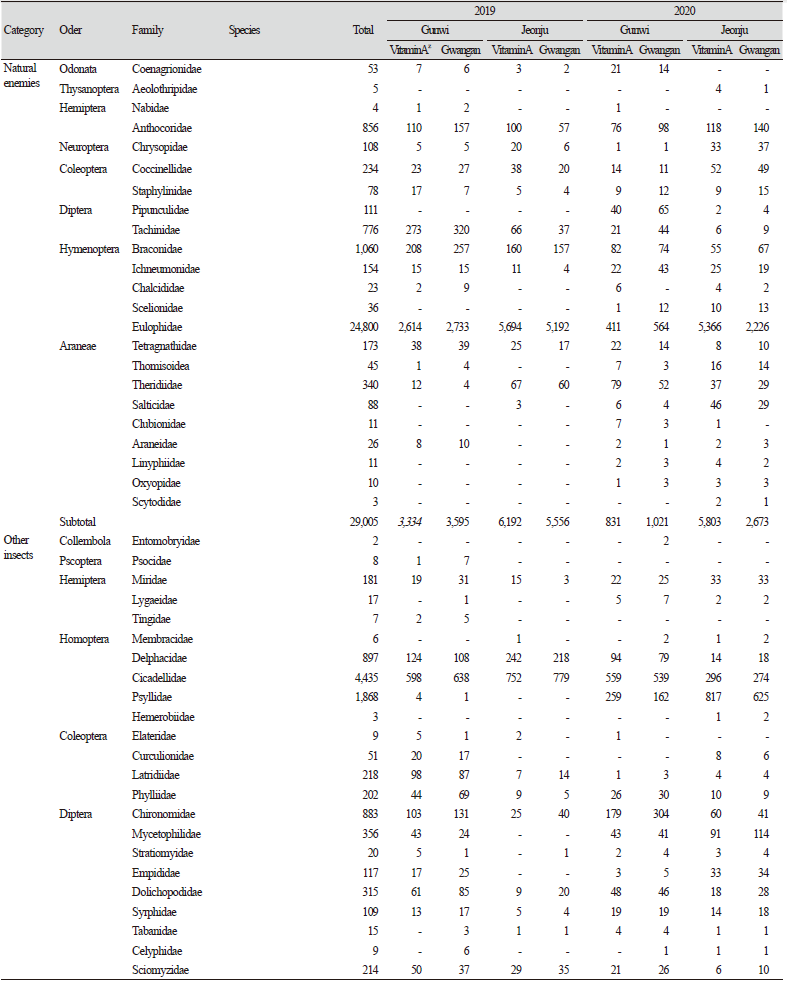
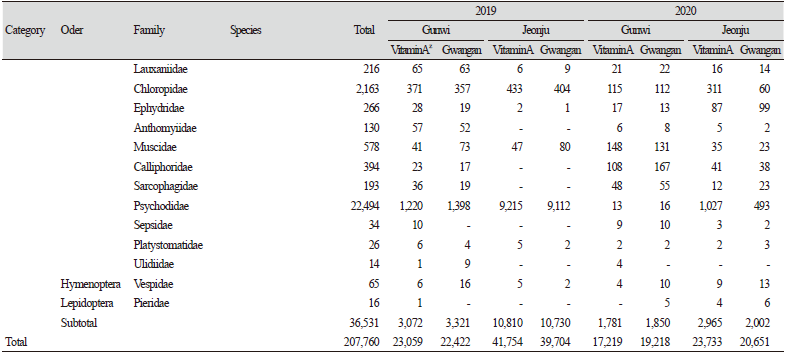
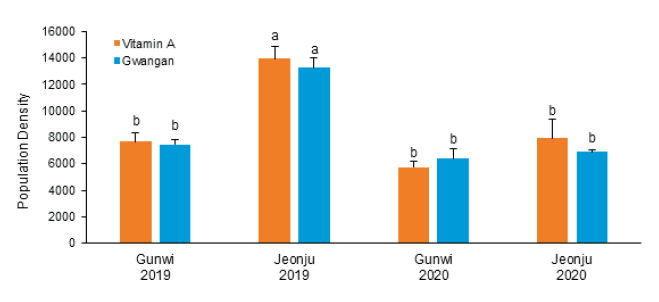
Fig. 1. Average population densities in common plant dwelling insects and arachnids captured using yellow sticky trap at fields planted with two different genotypes of soybean at Gunwi and Jeonju, Korea in 2019 and 2020. The results shown are the mean ± SD, n = 3 replicates for each group. Vitamin A: Vitamin A-enhanced transgenic soybean. a, b: The least significant difference (LSD) test at p < 0.05.
기능군별 발생현황
조사기간 동안 두 지역에서 채집된 곤충들을 기능군으로 구분하여 분석한 결과, 해충류가 68.46%, 천적류가 13.96%, 기타곤충이 17.58%로, 해충의 발생밀도가 가장 높았다. 채집된 곤충 중 해충군에서는 꽃노랑총채벌레, 대만총채벌레가 속해있는 총채벌레과(Thripidae)의 발생량이 68.3%로 대부분을 차지하였으며 천적군은 좀벌과(Eulophidae)와 고치벌과(Braconidae) 등의 기생성 벌목이 12.5%로 우점하였다. 꼬마거미과(Theridiidae), 갈거미과(Tetragnathidae) 등의 거미류(Araneae) 또한 0.3%의 발생량을 보였다. 기타 곤충군에서는 나방파리과(Psychodidae), 노랑굴파리과(Chloropidae) 등 파리목이 13.7%로 대부분을 차지하였으며 매미충과(Cicadellidae), 나무이과(Psyllidae) 등 매미목(Homoptera) 곤충류 또한 3.5%의 발생량을 보였다(Fig. 2).
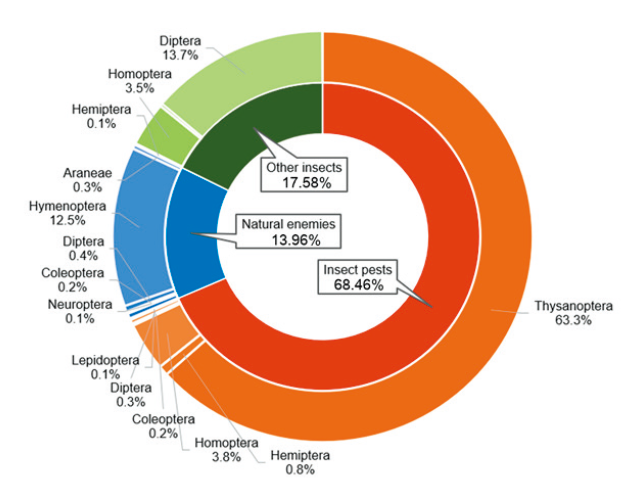
전체 개체군 중 해충군은 총채벌레과, 진딧물류, 매미충류 등을 포함한 매미목, 노린재목(Hemiptera), 딱정벌레목(Coleoptera) 등을 포함하여 총 7목 27과 43종의 곤충류가 발생하였다(Table 1 and 2). 2019년 군위에서는 총 28종의 해충이 조사되었으며, 총채벌레목이 92.12% (비타민A 강화콩 5,160.7 ± 646.4개체, 광안콩 4,714.3 ± 263.6개체)로 우점하였고, 매미목은 전체의 약 6.50%를 차지하였다. 다음으로 노린재목 0.84%, 딱정벌레목 0.34%, 나비목(Lepidoptera)이 0.20%, 그리고 메뚜기목이 0.01%를 차지하였다. 비타민A 강화콩과 광안콩에서 해충 발생 양상은 유사하였으며, 개체군밀도에서도 뚜렷한 통계적 유의차는 보이지 않았다. 2019년 전주에서는 총 22종의 해충이 조사되었으며, 총체벌레목이 92.53% (비타민A 강화콩7,635.0 ± 1024.7개체, 광안콩 7,222.3 ± 513.7개체)로 가장 우점하였으며, 다음으로 매미목 6.16%, 노린재목 1.06%, 나비목이 0.12%, 딱정벌레목이 0.11% 순이었다. 전주에서는 군위에서와 달리 딱정벌레목의 발생량이 다소 낮았으며, 다른 해충의 발생은 상대적으로 높았다. 또한 전체적인 해충 발생량은 전주가 군위보다 더 많은 등 지역간 차이가 나타났다. 그러나, 비타민A 강화콩과 광안콩에서의 해충 발생양상은 유사한 경향을 보였으며 개체군밀도는 역시 통계적 유의성이 없었다. 따라서 비타민A 강화콩이 광안콩의 해충 발생에는 특별한 영향을 미치지는 않는 것으로 판단된다. 2020년 군위에서는 총 31종의 해충이 조사되었으며, 이들 중 총채벌레목이 92.67% (비타민A 강화콩 4,511.3 ± 445.7개체, 광안콩 5,050.3 ± 568.1개체)로 가장 우점하며 해충의 대부분을 차지하였다. 매미목은 3.82%, 노린재목은 1.64%, 파리목 1.51%, 딱정벌레목 0.25%, 나비목 0.09%, 메뚜기목 0.01% 순으로 차지하였다. 또한, 2019년 군위와는 달리 파리목 해충이 새롭게 조사되는 등 조사연도 간의 차이가 나타났다. 하지만 비타민A 강화콩과 광안콩의 해충 발생양상은 유사하였으며 개체군밀도에서도 통계적 유의성은 나타나지 않았다. 2020년 전주에서는 총 36종의 해충이 조사되었으며, 이들 중 총채벌레목이 92.56% (비타민A 강화콩 4,642.7 ± 786.5개체, 광안콩 4,903.7 ± 923.4개체)로 가장 우점하며 해충의 대부분을 차지하였다. 또한, 매미목 5.32%, 노린재목 1.04%, 파리목 0.65%, 딱정벌레목 0.31%, 나비목 0.11%, 메뚜기목 0.01%를 차지했으며 다른 해충의 발생은 상대적으로 매우 낮았다. 총채벌레목, 노린재목, 파리목은 군위에 비해 전주에서 발생량이 적은 반면, 매미목, 딱정벌레목은 상대적으로 전주에서 발생량이 더 많은 등 지역간 차이가 나타났다. 또한 2019년 전주와는 달리 파리목 해충이 새롭게 조사되었고 전체 해충 발생량이 크게 감소하는 등 조사연도간의 차이가 보였다. 그러나 비타민A 강화콩과 광안콩에서 해충 발생양상은 유사하였으며, 개체군밀도에서도 통계적 유의성이 나타나지 않았다(Table 2). 따라서 비타민A 강화콩과 non-GM 콩인 광안콩에서 해충 발생 정도는 조사연도 및 지역 간에서 차이가 나타났으나, 동일 조사연도 및 지역 내에서는 유사한 경향을 보였기 때문에 비타민A 강화콩이 해충 발생에 특별한 영향을 미치지는 않는 것으로 판단된다.
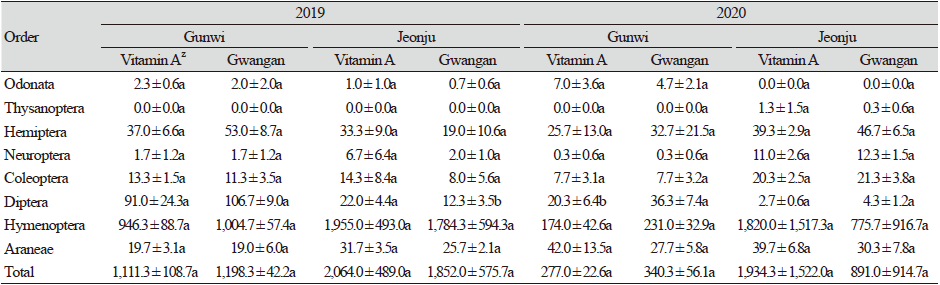
전체 개채군 중 천적군은 좀벌, 고치벌, 기생파리 등 기생성 천적곤충류와 꽃노린재, 무당벌레, 풀잠자리와 같은 포식성 천적곤충류와 거미류를 포함하여 총 8목 23과가 조사되었다(Table 1 and 3). 2019년 군위에서는 총 7목 15과의 천적곤충류와 거미류가 조사되었으며, 이들 중 좀벌과, 고치벌과 등을 포함하는 벌목이 84.48% (비타민A 강화콩 946.3 ± 88.7개체, 광안콩 1,004.7 ± 57.4개체)로 가장 우점하였으며, 파리목이 8.56%, 노린재목 3.88%, 거미류 1.67%, 딱정벌레목 1.07%, 잠자리목(Odonata) 0.19%, 풀잠자리목 0.14% 순으로 나타났다. 비타민A 강화콩과 광안콩에서 천적류 발생양상은 유사하였으며, 개체군밀도에서도 통계적 유의성은 나타나지 않았다. 전주에서는 총 7목 12과의 천적곤충 및 거미류가 조사되었으며, 이들 중 벌목이 95.49% (비타민A 강화콩 1,955.0 ± 493.0개체, 광안콩 1,784.3 ± 594.3개체)로 가장 우점하였으며, 거미류 1.46%, 노린재목 1.34%, 파리목 0.88%, 딱정벌레목 0.57%, 풀잠자리목 0.22%, 잠자리목 0.04% 순으로 나타났다. 전주에서는 군위에 비해 적은 비율의 파리목, 매미목이 채집되는 등 지역간의 차이가 보였다. 하지만 비타민A 강화콩과 광안콩에서 천적류 발생양상은 유사하였다. 개체군밀도는 파리목에서 통계적 유의성이 나타났지만, 이는 적은 개체수로 인해 약간의 차이에도 유의성이 나타나는 것으로 판단된다. 또한 나머지 천적곤충류 및 거미류에서의 통계적 유의성 또한 없는 점으로 미루어 보아 비타민A 강화콩이 천적류 발생에 특별한 영향을 미치지 않은 것으로 판단된다. 2020년 군위에서는 총 7목 21과의 천적곤충류 및 거미류가 조사되었으며, 이들 중 벌목이 65.60% (비타민A 강화콩174.0 ± 42.6개체, 광안콩 231.0 ± 32.9개체)로 우점하였으며, 거미류 11.29%, 노린재목 9.45%, 파리목 9.18%, 딱정벌레목 2.48%, 잠자리목 1.89%, 풀잠자리목 0.11% 순으로 나타났다. 2019년 군위보다 2020년 군위에서 천적류 발생량에 비해 더 많은 거미류가 잡히는 등 조사연도별 차이가 나타났다. 그러나 비타민A 강화콩과 광안콩에서 천적류 발생양상은 유사하였다. 개체군밀도에서는 파리목에서 통계적 유의성이 나타났지만, 이 역시 적은 개체수로 인해 약간의 차이에도 유의성이 나타나는 것으로 판단된다. 또한 파리목을 제외한 나머지 천적곤충류 및 거미류에서는 통계적 유의성이 나타나지 않은 것으로 미루어 보아 비타민A 강화콩이 천적류 발생에 특별한 영향을 미치지 않은 것으로 판단된다. 전주에서는 총 7목 21과의 천적곤충류 및 거미류가 조사되었으며, 벌목이 91.87% (비타민A 강화콩 1,820.0 ± 1,517.3개체, 광안콩 775.7 ± 916.7개체)로 우점하였으며, 노린재목 3.04%, 거미류 2.48%, 딱정벌레목 1.47%, 풀잠자리목 0.83%, 파리목 0.25%, 총채벌레목 0.06% 순으로 나타났다. 군위에서는 잠자리목이 채집되었고 총채벌레목은 채집되지 않았으며, 전주에서는 잠자리목이 채집되지 않았고 총채벌레목이 채집되는 등 지역간 차이를 보였다. 또한 2019년 전주에서보다 2020년 전주에서 더 많은 노린재목이 채집되는 등 조사연도간의 차이 역시 나타났다. 그러나 비타민A 강화콩과 광안콩에서 천적류 발생양상은 유사하였으며, 개체군밀도에서 통계적 유의성은 나타나지 않았다(Table 3). 따라서 비타민A 강화콩과 non-GM 콩인 광안콩에서 천적류 발생은 조사연도 및 지역 간에서 차이가 나타났으나, 동일 조사연도 및 지역 내에서는 유사한 경향을 보였기 때문에 비타민A 광안콩이 천적류에 특별한 영향을 미치지는 않는 것으로 판단된다.
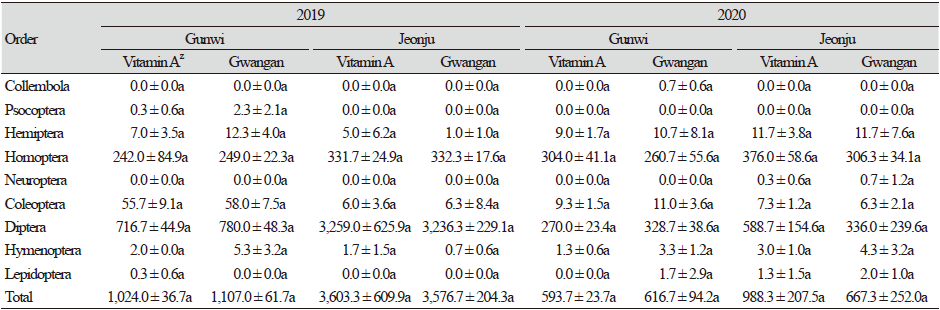
조사기간 동안 해충과 천적군을 제외한 기타 곤충군은 나방파리과, 매미충과, 노랑굴파리과를 포함해 총 8목 36과가 조사되었다(Table 1 and 4). 2019년 군위에서는 총 7목 33과의 곤충류가 조사되었으며, 이들 중 나방파리과, 노랑굴파리과, 깔따구과 등 파리목이 70.23% (비타민A 강화콩 716.7 ± 44.9개체, 광안콩 780.0 ± 48.3개체)로 가장 우점하였으며, 멸구과, 매미충과 등 매미목이 23.04%, 섶벌레과, 잎벌레과 등이 포함된 딱정벌레목은 5.33%, 노린재목 0.91%, 벌목 0.34%, 다듬이벌레목(Pscoptera) 0.13%, 나비목 0.02%를 차지하였다. 비타민A 강화콩과 광안콩에서 해충 및 천적을 제외한 기타곤충류 발생양상은 유사하였으며, 개체군 밀도에서도 통계적 유의성은 나타나지 않았다. 전주에서는 총 5목 20과의 곤충류가 조사되었으며, 파리목이 90.46% (비타민A 강화콩 3,259.0 ± 625.9개체, 광안콩 3,236.3 ± 229.1개체)로 대부분을 차지하였고, 매미목이 9.25%로 나타났다. 그 외 딱정벌레목 0.17%, 노린재목 0.08%, 벌목 0.03% 순으로 나타났다. 전주에서는 군위와 달리 다듬이벌레목이 채집되지 않았고 채집된 기타곤충류 중 파리목이 상대적으로 더 많은 비율을 차지하는 등 지역간 차이를 보였다. 그러나 비타민A 강화콩과 광안콩에서 해충 및 천적을 제외한 기타곤충류 발생양상은 유사하였으며, 개체군밀도에서도 뚜렷한 통계적 유의성은 보이지 않았다. 2020년 군위에서는 총 7목 32과의 곤충류가 조사되었으며, 파리목 49.5% (비타민A 강화콩 270.0 ± 23.4개체, 광안콩 328.7 ± 38.6개체), 매미목이 46.7% (비타민A 강화콩 304.0 ± 41.1개체, 광안콩 260.7 ± 55.6개체)로 기타곤충류의 대부분을 차지하였으며, 딱정벌레목 1.68%, 노린재목 1.62%, 벌목 0.39%, 나비목 0.14%, 톡토기목(Collembola) 0.06%를 차지하였다. 비타민A 강화콩과 광안콩에서의 기타곤충류 발생양상은 유사하였으며, 개체군밀도에서도 뚜렷한 통계적 유의성은 없었다. 전주에서는 총 7목 31과의 곤충류가 조사되었으며, 파리목이 55.85% (비타민A 강화콩 588.7 ± 154.6개체, 광안콩 336.0 ± 239.6개체), 매미목이 41.21% (비타민A 강화콩 376.0 ± 58.6개체, 광안콩 306.3 ± 34.1개체)로 기타곤충류의 대부분을 차지하였으며 노린재목 1.41%, 딱정벌레목 0.83%, 벌목 0.44%, 나비목 0.20%, 풀잠자리목 0.06% 순으로 나타났다. 군위에서 톡토기목이 채집되었고 풀잠자리목이 채집되지 않은 반면, 전주에서는 반대로 톡토기목이 채집되지 않았고 풀잠자리목이 채집되는 등 지역간 차이를 보였다. 또한 군위와 전주 모두 2019년에 비해 2020년에 매미목이 훨씬 높은 비율로 채집되는 등 조사연도 간의 차이를 보였다. 그러나 비타민A 강화콩과 광안콩에서 기타곤충류 발생양상은 유사하였으며, 개체군밀도에서도 뚜렷한 통계적 유의성은 없었다(Table 4). 따라서 비타민A 강화콩과 non-GM 콩인 광안콩에서 기타곤충류의 발생은 조사연도 및 지역 간에서 통계적 차이를 보였으나, 동일 조사연도 및 지역 내에서는 유사한 경향을 보였기 때문에 비타민A 강화콩이 기타곤충류 발생에 특별한 영향을 미치지는 않는 것으로 판단된다.
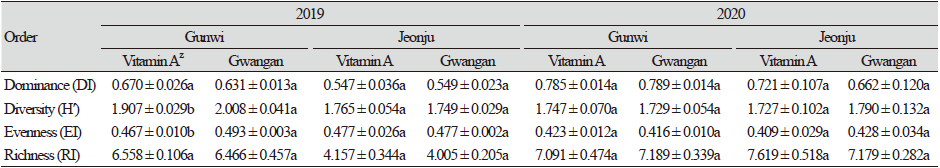
2019년과 2020년에 군위와 전주의 비타민A 강화콩과 광안콩의 재배지에서 채집된 곤충상의 군집의 종다양성을 분석한 결과, 전체 채집된 곤충에서 우점도 지수(DI)는 군위에서 0.631 - 0.789, 전주에서 0.547 - 0.721로 품종간 우점도 지수는 통계적 유의성을 나타내지는 않았다. 우점도지수가 높게 나온 이유는 꽃노랑총채벌레와 대만총채벌레를 포함하는 총채벌레목의 개체수가 다른 과에 비해 많이 채집되었기 때문이라고 판단된다. 다양도 지수(H’)는 군위에서 1.729 - 2.008, 전주에서 1.727 - 1.790로 군위에서 다소 높아 통계적인 차이를 보였으며, 2019년 군위에서는 비타민A 강화콩에서 1.907, 광안콩에서 2.008로 품종간 통계적 유의성이 확인되었다. 이는 적은 개체수가 잡힌 분류군으로 인해 약간의 차이에도 유의성이 나타나는 것으로 판단되며, 2019년 전주 및 2020년 군위와 전주에서는 그 차이가 나타나지 않는 것으로 보아 비타민A 강화콩이 다양도 지수에 특별한 영향을 미치지 않은 것으로 판단된다. 균등도 지수(EI)는 군위에서 0.416 - 0.493, 전주에서 0.409 - 0.477로 분석되어 전주에 비해 군위에서 조금 더 다양한 종들이 채집되었으며, 2019년 군위에서는 비타민A 강화콩에서 0.467, 광안콩에서 0.493으로 품종간 틍계적 차이가 나타났다. 이 역시 적은 개체수가 잡힌 분류군으로 인한 것으로 판단되며, 2019년 전주 및 2020년 군위와 전주에서는 그 차이가 나타나지 않는 것으로 보아 비타민A 강화콩이 균등도 지수에 특별한 영향을 미치지 않은 것으로 판단된다. 종풍부도 지수(RI)는 군위와 전주 두 지역 모두 2019년에 비해 2020년의 종풍부도가 높아진 결과를 보였다. 품종별로는 통계적인 유의성은 보이지 않았다(Table 5). 따라서 지역과 연도간에는 종 다양성의 차이를 보였으나, 동일 조사연도 및 지역 내에서는 뚜렷한 차이를 보이지 않았으며, 비타민A 강화콩이 재배지 내의 곤충 종다양성에 특별한 영향을 주지 않는 것으로 판단된다. 이러한 결과는 곤충상 조사방법의 차이가 있지만, 2015년에 수행된 비타민E 강화콩 재배지에서 발생하는 곤충상과 생물다양성 비교 분석 결과와 유사하였다(Oh et al., 2017). 또한, 해충저항성 GM 벼 재배지의 해충 및 천적류에 대한 조사에서도, 본 연구와 비슷한 결과를 보였다(Amin et al., 2020b).
주성분분석
LMO 격리포장(전주, 군위)에서 2년(2019, 2020) 동안 황색 끈끈이 트랩을 이용하여 확보한 곤충상 데이터를 통계분석하여 발생 양상에 변화를 보인 곤충이 무엇인지 확인하기 위해 다변량분석법 중 PCA와 OPLS-DA를 수행하였다. PCA는 복잡한 데이터 집합에서 특징적인 정보를 추출할 때 유용하다(Park et al., 2013; Amin et al., 2020b). PCA 결과, 비타민A 강화콩을 재배하였을 때 발생하는 곤충상은 모품종인 광안콩과 비교하였을 때 분석한 모든 데이터 세트에서 확연한 차이는 나타나지 않았다. 하지만, 재배 연도와 지역에 따라서는 곤충상의 차이가 명백하게 보여졌다(Fig. 3). 2020년 군위의 비타민A 강화콩 샘플 중 하나(Fig. 3A)에서 점선 타원 안에 속하지 않은 샘플)가 2020년 군위와 2019년 군위 샘플들 중간에 위치하지만, 이는 제1 주성분과 제2 주성분에 의해(총 데이터 변이의 41.2% 차지) 전주 지역과 2019년 군위의 곤충상과 분리되었다. Score plot에서 보여지는 샘플간 곤충상의 차이를 좀 더 자세히 알아보기 위해 loding plot을 확인한 결과, 세 그룹(해충군 및 천적군, 기타 곤충군) 내에서 특이적으로 발생의 차이를 보인 곤충군은 없었다(Fig. 3B).
재배 연도와 지역에 따라서 샘플의 구분을 최적화하고 그 차이를 분석하기 위해 OPLS-DA를 수행하였다. OPLS-DA는 두개의 임의적 그룹 지정을 통해 샘플 간의 최대한의 분리를 나타내기 위해 사용되는 분석법이다(Kim et al., 2020b). 그룹을 재배 연도와 지역으로 각각 지정하여 OPLS-DA를 분석한 결과, 곤충상은 연도(2019, 2020)와 지역(군위, 전주)으로 명백히 구분되었다(Fig. 3C and 3E). 이러한 그룹화에 영향력을 미친 곤충이 무엇인지 확인하기 위해 VIP값을 확인하였다. VIP는 모델에 대한 기여도에 따라 변수의 순위를 지정하는 것으로 값이 1.0 보다 크면 샘플의 그룹을 형성하는데 영향력이 큼을 의미한다(Lee et al., 2017). 재배 연도별로 그룹화하는데 있어서는, 거미류를 포함한 곤충류 86개 과 중 34개 과의 VIP값이 1을 넘었으며 18과는 VIP값이 1.5를 넘었다. 이 18개 과 중에는 기타 곤충류가 50%를 차지하며 가장 많았으며 그 다음으로는 해충과 천적 순으로 나타났다(Fig. 3D). 재배 지역별로 그룹화하는데 있어서는, 거미류를 포함한 곤충류 37개 과의 VIP값이 1을 넘었으며 그 중 16개 과가 VIP값이 1.5를 넘었다. 기타 곤충류가 56.4%를 차지하며 가장 많았으며 그 다음으로는 천적과 해충 순으로 나타났다(Fig. 3F). 재배 연도와 지역별로 곤충상을 분류하는데 있어 19개 과의 곤충이 1이 넘는 VIP값을 보여주었다. 본 연구에서 수행한 다변량분석을 통해, 콩의 유전 형질은 식물이 재배된 곳의 곤충상 변화에 영향을 미치지 않으며, 재배 연도와 지역에 따라 곤충상의 변화가 더 큼을 확인할 수 있었다.
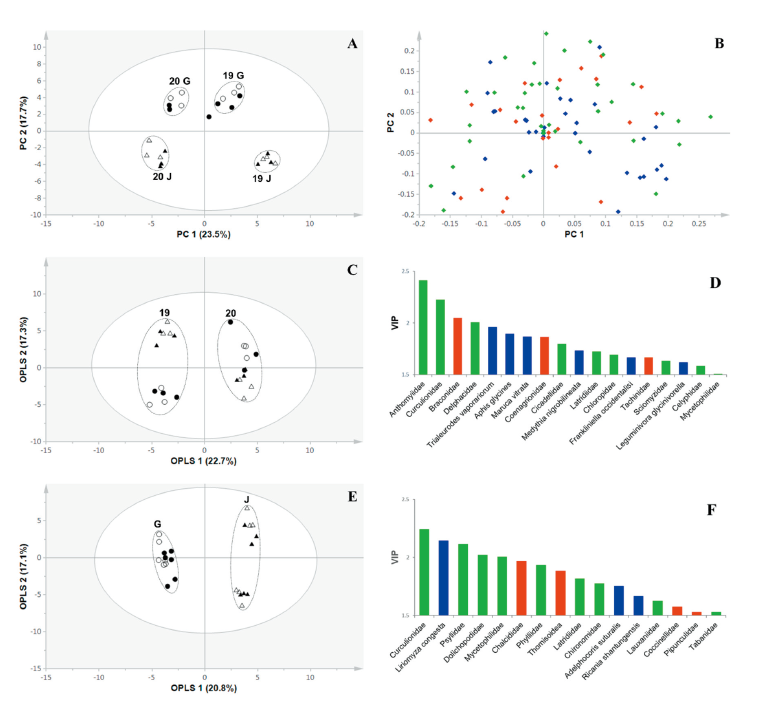
Fig. 3. Score (A) and loading plots (B) of the principal component analysis (PCA) results obtained from data of non-target insects and arachnids captured using yellow trap at fields planted with two different genotypes of soybean in two years at two different regions. Score plots and variable importance in the prediction (VIP) values of the orthogonal partial least squares-discriminant analysis (OPLS-DA) results obtained from data of the PCA results by cultivation year grouping (C and D) and cultivation region grouping (E and F). Circle, Gunwi; triangle, Jeonju; empty symbol, Gwangan; black-filled symbol, vitamin A enhanced soybean; 19, 2019; 20, 2020; G, Gunwi; J, Jeonju; blue, petss; red, natural enemies; green, other insects.
Conclusion
본 연구에서는 거미류를 포함한 곤충류의 다양도 및 풍부도에 대해 비타민A 강화콩의 영향을 평가하기 위해 포장 재배 조건 하에 시험하였다. 비타민A 강화콩은 카로티노이드 함량을 증가시키기 위해 카로티노이드 생합성 유전자인 Psy와 CrtI를 발현시켰다. 2019년과 2020년 2년간 경북대학교 군위 LMO 격리포장과 국립농업과학원 전주 LMO 격리포장에서 비타민A 강화콩과 non-GM 콩인 광안콩을 비교하였다. 조사기간 총 13목 81과 207,760개체의 곤충류 및 거미류가 채집되었다. 품종별로는 비타민A 강화콩 재배지에서 105,765개체, 광안콩 재배지에서 101,995개체가 조사되었다. 연구 전반에 걸쳐 곤충상의 변이에 대해 유의한 차이는 나타나지 않았다(p <0.05). 통계분석 결과, 우점도 지수(DI), 종풍부도 지수(RI)에서는 품종, 지역간 차이가 없었고, 다양도 지수(H'), 균등도 지수(EI)에서는 2019년 군위를 제외한 다른 곳에서 통계적 유의차를 나타내지 않았다. 본 연구를 통해 얻은 곤충상 데이터를 이용하여 주성분분석(PCA)과 직교부분최소자승판별분석(OPLS-DA)을 수행한 결과, 비타민A 강화콩과 광안콩 간의 곤충상에 차이가 없으며, 재배 연도와 지역을 기준으로 곤충상의 차이가 나타나는 것을 볼 때, 비타민A 강화콩이 재배지의 곤충상 변화에 영향을 미치지 않음을 확인할 수 있었다.
Authors Information
Sung-Dug Oh, https://orcid.org/0000-0001-8574-6773
Kihun Ha, https://orcid.org/0000-0002-2938-1906
Soo-Yun Park, https://orcid.org/0000-0002-2030-6440
Seong-Kon Lee, https://orcid.org/0000-0002-0939-9391
Do won Yun, https://orcid.org/0000-0002-6633-5685
Kijong Lee, https://orcid.org/0000-0003-3111-322X
Sang Jae Suh, https://orcid.org/0000-0002-7489-3193

