Introduction
국가 간 교류가 활발해지면서 마약류 및 불법 식품 및 의약품의 국내 반입과 테러와 같은 새로운 위협요소들이 전 세계적으로 상존하고 있고, 이로부터 국민의 안전을 지키는 국경 안보가 새롭게 이슈로 부각되고 있다. 이러한 위협요소들을 사전에 탐지하는 특수 목적용 반려견의 활동이 매우 중요해지고 있다. 현재 우리나라는 국민 안전을 위해 국방부와 경찰청에서는 폭발물
탐지견을, 농림축산식품부에서는 검역 탐지견, 관세청에서는 마약 탐지견을 그리고 소방방재청에서는 인명 구조견을 각각 양성하고 운용 중에 있다. 특수목적 임무를 수행하는 반려견을 양성하고 생산하는데 일반견을 이용할 경우 양성율은 20 - 30% 수준으로 알려져 있으며, 비용은 연간 약 1억 3천 만원으로 많은 비용과 시간이 소요된다. 따라서, 첨단기술인 체세포복제기술을 이용하여 특수목적 업무에 적합한 우수한 개체를 복제하여, 보급하는 기술이 개발 되었다. 그러나 우수한 개체를 확보하여 단기간에 특수목적 업무를 하는 반려견을 양성하고 현장에 투입하는데 있어서 장애요인은 근골격계 질환이 발생하여 특수 업무를 방해 한다는데 있다. 이러한 유전적 장애요인을 사전에 예측하고, 유전적으로 질병 소인을 가지고 있는 개체를 진단하는 기술이 개발 되어야 한다. 이러한 유전적 장애요인과 연관된 유전자를 탐색하기 위해서는 전장유전체 연관분석(Genome wide association)이라는 기술을 사용하고 있는데, 이는 유전적 장애요인을 유발하는 원인 유전자(causal mutation)와 연관불평형(linkage disequilibrium)관계에 있는 유전 마커를 탐색하는 기술로 개체의 유전 질환과 연관된 표현형과 유전자형 간 통계적인 연관성을 탐색한다(Vonholdt et al., 2010).
연관불평형(linkage disequilibrium, LD)은 종별로 매우 상이하며, 이러한 상이한 유전체의 연관불평형이 개(dog), 고양이(cat), 양(sheep), 그리고 소(cattle)에서 보고 되었다(Pritchard and Przeworski, 2001; Khatkar et al., 2008; Gray et al., 2009; Vonholdt et al., 2010; García-Gámez et al., 2012; Alhaddad et al., 2013). 최근 순계품종 반려견(purebreed dog) 연구에서 보여주는 것과 같이 연관불평형 정보는 사람의 것보다 훨씬 크다는 것이 보고 되었다(Sutter et al., 2004; Vonholdt et al, 2010). Parker (2012)의 연구에 따르면, 순계 품종 반려견 집단에서 집단의 유전적 bottleneck이 발생하고 있으며 공통선조효과(founder effect)에 의해서 예상한 것 보다 훨씬 큰 연관불평형구조가 형성되어 있다고 보고 하였다. 특히 반려견의 골격계 질병 중에서 중증이라고 할 수 있는 고관절 이형성증(Canine Hip Dysplasia, CHD)질환은 70년 전에 처음 보고 되었으며, 이는 혈통 내에서 교배하는 근친 교배에 의하여 발생하며, 유전적으로는 여러 개의 유전자에 의한 다인성으로 보고 되고 있다(Schnelle, 1935).
따라서, 본 연구의 목적은 국내 특수목적을 수행하는 대형종 반려견 집단의 유전적 특성을 알아보기 위하여 수행되었다. 대표적으로 세퍼드, 말리노이즈, 그리고 리트리버 품종의 연관불평형 및 유효집단크기 및 다른 품종과 유전적 다양성을 분석함으로 국내 특수목적견의 유전적 특성에 기초한 지속 가능한 집단을 유지하기 위하여 수행하였다.
Materials and Methods
공시 재료
본 연구는 농촌진흥청 국립축산과학원 및 국방부 육군 군견대의 동물복지위원회(Animal Care and Use Committees)에서 제시한 가이드라인을 준수하여 표본을 추출 하였다. 국방부 육군 군견대에서 보유하고 있는 특수목적 임무를 위하여 훈련중인 세퍼드(n = 57) 및 말리노이즈(n = 12)로부터 경정맥 채혈을 통하여 시료를 확보 하였다. 이와 함께 두 품종과 비교하기 위하여 공공 데이터베이스로부터 확보한 리트리버(n = 723) 말리노이즈(n = 6) 그리고 세퍼드(n = 284)를 이용하였다.
gDNA 추출 및 유전자형 결정
경정맥 채혈을 통해 확보한 전혈로부터 genomic DNA는 표준 프로토콜(Meurs et al., 2010)을 이용하여 추출 하였다. 세 품종에 대한 대용량 유전자형을 결정하기 위하여 Illumina CanineHD BeadChip (Illumina, San Diego, CA, USA)을 이용하여 유전자형을 결정하여 총 170,000개의 단일염기변이(single nucleotide polymorphism; SNP)를 유전자형 결정 하였다.
SNP 품질 관리(quality control)
대용량 유전자형에 대한 표준화 정도 관리(quality control)는 PLINK1.7 (S. Purcell, http://pngu.mgh.harvard.edu/ purcell/plink/) 를 이용하여 수행 하였다(Purcell et al., 2007). 먼저 missing genotype은 10% 미만을 제거 하였고, call rate 95%로 설정 하였다. 아울러, 유전자형의 minor allele frequency (MAF)는 0.05 미만으로 설정하여, 이에 미치지 못한 SNP는 제거 되었고, Hardy-Weinberg equilibrium (HWE)은 0.00005 미만으로 설정하였다.
연관불평형(Linkage disequilibrium) 분석 및 유효집단(effective population size) 크기 분석
세 개의 반려견 집단에서 연관불평형의 크기는 서로 다른 두 대립유전자가 서로 연관되어 유전되는 정도를 나타내는 값으로 먼저 D값을 구한 후에 이를 표준화된 D’을 구하고, 이를 이용하여 두 대립유전자간의 상관을 보여주는 통계량인 r2 값을 추정하였다(Lewontin et al., 1964; Hill et al., 1968; Lee et al., 2013). 두 개의 마커 간 연관 불평형을 계산하기 위하여 Plink1.7의 --ld 기능을 이용하여 분석하였다. 두 마커 간 거리를 나타내는 distance bin은 0 - 10; > 10 - 20; > 20 - 40에서 최대 > 20,000 - 50,000로 계산 하였다. 동일 염색체에 존재하는 두 마커 좌위 A, B의 연관불평형 크기(r2)는 다음의 수식을 이용하여 계산 하였다.
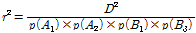 (1)
(1)
여기서 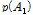 과
과 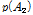 그리고
그리고 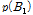 과
과 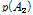 는 두 대립유전자의 빈도를 나타내며, D는 다음의 수식을 이용하여 계산 하였다.
는 두 대립유전자의 빈도를 나타내며, D는 다음의 수식을 이용하여 계산 하였다.
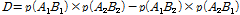 (2)
(2)
여기서, 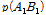 과
과 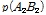 그리고
그리고 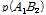 과
과 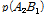 는 두 마커 간 일배체형의 빈도를 나타낸다.
는 두 마커 간 일배체형의 빈도를 나타낸다.
유효집단크기 추정에 있어서 연관불평형 정보는 매우 중요하게 이용되는 통계량으로 연관불평형은 재조합 및 매 세대 선발에 의한 돌연변이의 누적 효과에 의해서 발생된다. 즉, 재조합의 경우 두 마커 간 거리가 먼 좌위에서는 발생할 확률이 높고, 매우 가까운 위치에서는 발생할 확률이 매우 낮다. 따라서 두 마커 간 거리와 연관불평형 값(r2)을 추정한다면, 돌연변이가 없다는 가정하에 Sved (1971)가 제한한 아래의 식에 의해 유효집단 크기를 추정 할 수 있다. 따라서, 본 연구에서 유효집단 크기는 아래와 같이 추정하였다(Lee et al., 2015).
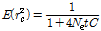 (3)
(3)
여기서 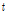 는 이전 세대,
는 이전 세대, 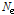 는 유효집단 크기, 그리고
는 유효집단 크기, 그리고 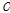 는 Morgan 단위로 표현되는 마커의 거리이며, 세대별,
는 Morgan 단위로 표현되는 마커의 거리이며, 세대별, 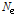 의
의
추정은 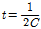 의 함수식으로 추정 하였다. 즉 두 마커 간 거리가 가까운 위치에서 발생한 연관불평형 값은 보다 오
의 함수식으로 추정 하였다. 즉 두 마커 간 거리가 가까운 위치에서 발생한 연관불평형 값은 보다 오
래 전 세대에서 발생한 것임을 알 수 있다. 위에 제시된 유효집단 크기는 함수식을 적용한 자체 스크립트를 제작하여 R package (The R project for statistical computing, ver.3.25; http://www.r-project.org)를 이용하여 분석 하였다.
이형 유전자 감쇠(Decay of Heterozygosity) 분석
특수 목적견의 이형 유전자 감쇠를 분석하기 위하여 plink software를 이용하여 분석한 유효집단 크기 분석 결과를 토대로 보존 유전학의 목적을 위해 이형 유전자의 변화를 계산하였다. 자연 선발과 인위 선발을 통하여 이형 접합 유전자가 감소하고 향후에 근친 교배를 예방하기 위하여 각 집단간 이형 유전자 감소를 면밀히 모니터링 해야한다. 계산은 다음과 같은 공식에 의하여 추정 하였다.
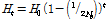 (4)
(4)
여기서 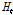 와
와 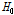 는 각각 시간
는 각각 시간  와 시간 0에서의 이형접합성이다
와 시간 0에서의 이형접합성이다
유전적 다양성(Genetic diversity) 분석
특수목적견의 유전적다양성을 분석하기 위하여 Plink software의 Multi-Dimensional Scale (MDS) 분석을 이용하여 집단간 유전적 다양성 분석을 수행하였다. 모든 상동염색체상의 SNP들을 이용하여 개체간의 identity-by-state (IBS) 거리를 측정한 후 IBS matrix를 R-Software를 이용하여 주성분 분석을 수행하여 개체간의 유전적 다양성을 계산 하였다.
Results and Discussion
순종의 개에서 유전 질환은 품종 내에서 강한 선발, 근친 교배 및 매우 제한된 수의 부모간 교배에 의한 유전적 격리로부터 발생한다(Mellersh, 2012). 이러한 형태의 개의 육종으로 전체 게놈영역에서 유전적 다양성이 급격하게 감소하여 열성 대립 형질에 대한 동형 접합이 높아져 질병의 유병률이 높아짐으로 반려견의 건강과 복지에 매우 큰 영향을 미친다(Rooney and Sargan, 2009). 반려견의 가축화 이후, 매우 다양한 형태로 인위적인 선발이 진행되어 왔으며, 이러한 인위적 선발이 고립된 개체군 또는 번식 개체군으로 이어져 열성 질병이 매우 흔하며 많은 품종이 특정 질환에 대한 위험이 증가한다(Ostrander, 2012). 그러므로 이러한 유전질환의 대비한 반려견의 육종은 소유주 및 육종가에게 중요한 관심사일 수밖에 없다. 현재 반려견의 유전질환에 대한 유전자 검사의 수가 증가하고 있고, 유전자검사 결과에 따라서 위험군의 개체를 육종집단에서 제거하고 있음에도 불구하고, 근원적인 해결책이 되지 않고 있다(Leroy and Abitbol, 2010). 그 이유는 유전질환을 조절하는 유전자가 하나가 아니라 여러 개이기 때문에, 집단에서 하나의 유전자변이검사를 통하여 위험군을 제거한다 하더라도 다른 유전자가 질병을 또다시 일으키게 된다. 따라서 현재 수행하고 있는 전략에 새로운 전략 즉, 현재 스크리닝 전략, 가계도 정보, EBV 또는 gEBV를 통합하는 전략은 유전적 다양성을 관리하는 동시에 유전된 질병의 수와 유행을 감소시킬 수 있는 대안이 된다(Farrell et al., 2015). 결과적으로 반려견 집단의 유전적 다양성, 근친 제어가 매우 중요한 요소이다.
국내 군견훈련소에서 운용하고 있는 품종 중에서 세퍼드와 말리노이즈를 대상으로 시료를 확보하였다. 이들 두 품종은 역사적으로 품종 정립이 잘 되어 있는 품종이며, 특히 세퍼드와 말리노이즈는 특수목적견인 군견으로서 매우 명성이 있는 품종으로 알려져 있다. 특수목적임무를 수행하는 군견을 중심으로 집단 유전학적 특성을 알아보기 위하여 기존에 shannon 등이 보고한 일반 집단으로부터 수집한 리트리버, 세퍼드 그리고 말리노이즈 품종을 더하여 군견의 품종과 이들 일반 집단의 품종 간 집단유전학적 특성을 분석하였다. 품종의 유전적 다양성 및 집단 유전학적 분석을 위하여 Canine 170 K chip을 이용하였으며, Table 1에서 보는 바와 같이 품종 별, 그리고 염색체 별 유전 변이의 수는 군견 세퍼드가 129,087개, 군견 말리노이즈가 115,861, 일반 집단 리트리버가 128,552, 일반 집단 세퍼드가 128,828개 그리고 일반 집단 말리노이즈가 120,212개로 분석되었다. 이러한 차이는 품종 간의 차이 또한 확보한 시료에 따라서 SNP QC결과가 다르게 나온 것으로 판단된다. 이들 단일염기변이(SNP)를 이용하여 총 5개 집단의 유전적 다양성을 분석하였다. Fig. 1에서 보는 바와 같이 개체 별 유전적 유사도를 보여주는 identity-by-decent (IBD)를 계산하여 이를 기반으로 5개 집단의 유전적 유사도를 주성분 분석(principle component)을 수행하였다. 군견에서 운용하는 말리노이즈와 일반 집단의 말리노이즈는 유전적으로 매우 유사하게 하나의 클러스터로 존재하고 있었고, 세퍼드의 경우도 일반 집단의 세퍼드와 군견 세퍼드가 같은 클러스터로 존재하고 있었다. 다만, 몇몇 군견 세퍼드의 경우 유전적으로 세퍼드와 일반 집단의 리트리버와 중간 정도의 유전적 유사도를 보였다. 이와 함께 커널 클럽의 리트리버는 유전적으로 매우 상이한 하나의 클러스터로 존재하고 있었다. 주성분 분석의 차원을 바꾸어 가면서 분석한 결과, 차원 2 (Dimension 2)와 차원 3 (Dimension 3)의 경우, 리트리버, 세퍼드와 말리노이즈가 매우 다른 유전적 유사성을 보였다. 일반 세퍼드 중 일부의 개체 3마리가 군견 세퍼드와 클러스터링 되고 나머지 세퍼드는 다른 군집으로 클러스터링 되는데 이는 일반 세퍼드와 군견 세퍼드에서의 유전적 차이를 나타내고 군견 세퍼드 보다 일반 세퍼드의 개체 수가 많아 군집 범위가 상대적으로 크게 나타났다. 아울러, 차원 3과 4의 경우 군견의 말리노이즈와 세퍼드의 경우, 커널 클럽의 말리노이즈 세퍼드와 상이한 유전적 유사성을 보였다.
|
Table 1. Number of SNPs used in this analyses after SNP quality control. 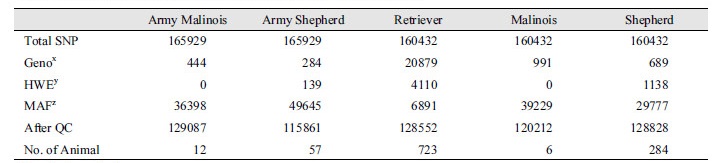
|
|
|
xGenotyping call rate. yHardy-Weinberg equilibrium. zGenotyping call rate. |
|
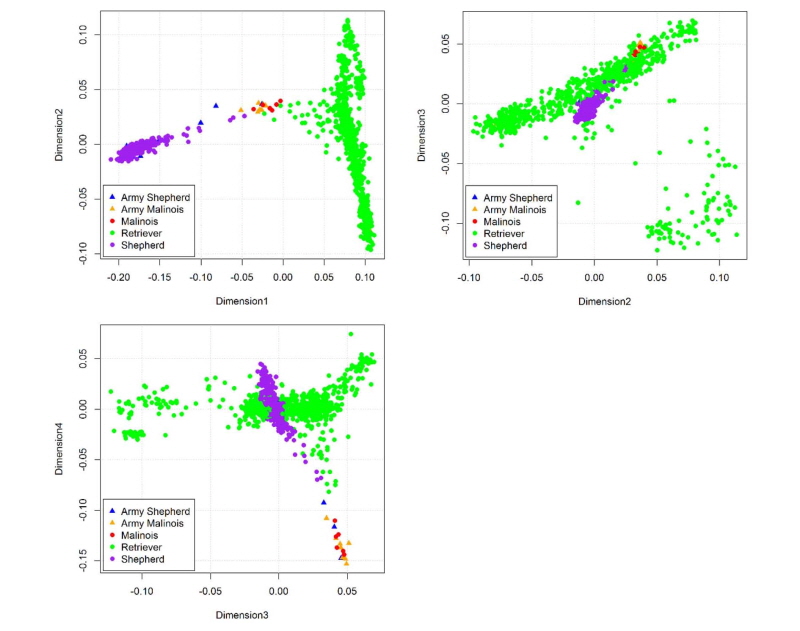
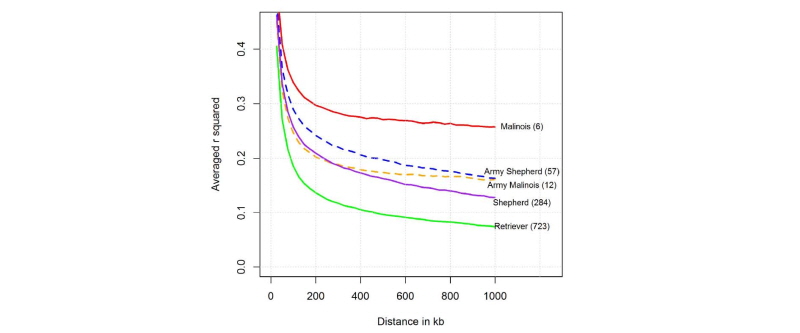
Fig. 2에서 보여주는 것과 같이, 본 연구에서 분석한 5품종에 대한 연관불평형분석 결과 총 6마리로 구성된 일반 말리노이즈가 마커와 마커 간 거리가 200 kb에서 약 0.3으로 매우 높은 결과를 보였고, 그 다음으로 군견 세퍼드(n = 57)가 0.25, 그리고 군견 말리노이즈(n = 12)가 0.2로 낮았고, 일반 집단 세퍼드(n = 284)가 200 kb를 기준으로 0.2로 분석되었고, 마커 간 거리가 커짐으로 지속적으로 작아지는 것을 보였다. 아울러 일반 집단 리트리버(n = 723)는 200 kb기준으로 0.13으로 분석 되었다. 즉 품종 간 연관불평형의 차이는 품종이 개량 되어지는 육종의 역사를 살펴볼 수 있는 매우 좋은 집단유전학적 모수이며, 본 연구에서 분석한 결과, 말리노이즈의 경우 군견과 일반 집단에서의 연관불평형 차이가 0.1로 분석되었고 세퍼드의 경우 군견과 일반 집단에서의 연관 불평형 차이가 0.05로 분석되어, 대체적으로 연구에 사용된 집단의 크기가 작을수록 분석결과의 편차가 크게 나타났다. 즉 일반 집단의 말리노이즈(n = 6)와 군견 말리노이즈(n = 12)는 품종의 특성도 있지만, 사용된 개체의 수가 매우 작아서 연관불평형 결과가 높게 나왔을 개연성도 있었다.
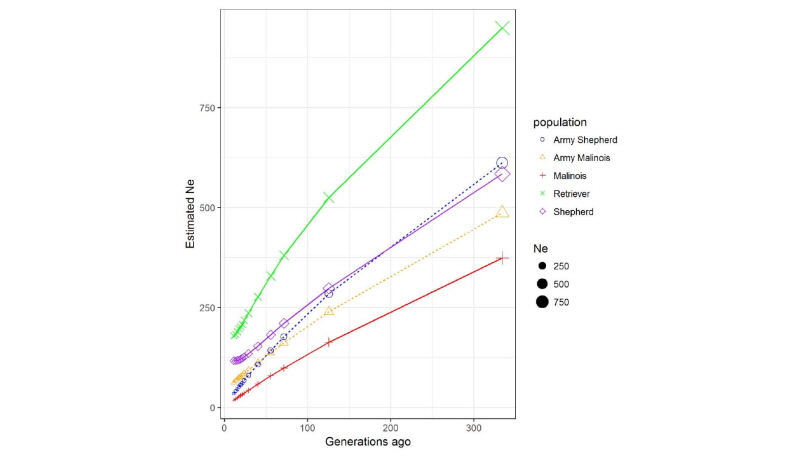
각 품종에서 분석된 연관불평형정보를 이용하여 개체의 유효 집단 크기를 분석하였다. 유효 집단의 크기(number of effective population size; Ne)는 “세대가 거듭되어도 집단내의 대립유전자의빈도가 변하지 않고 유지되는 최소 개체 수”라고 정의 할 수 있을 것이다(Hartl, 2000). 따라서 유효 집단의 크기는 근친을 제어하면서 지속 가능한 집단의 개량에 있어서 매우 중요한 지표일 것이다. Fig. 3과 Table 2에서 보여주는것과 같이, 약 11세대에 군견 말리노이즈의 유효 집단의 크기는 35두, 그리고 군견 세퍼드는 62두, 일반 집단 말리노이즈는 18두, 그러나 일반 집단 리트리버는 179두 그리고 일반 집단 세퍼드는 117두로 분석되었다. Choi et al. (2017) 에서 한국 개 품종과 여러 해외 개 품종을 이용하여 유효집단 크기 및 다양성에 대하여 수행된 결과에 따르면 17 개의 서로 다른 품종에서 평균 19개체의 샘플을 사용하였을 때의 평균 유효집단 크기는 본 연구에서 수행한 리트리버의 결과와는 유사하나 다른 품종에서는 상대적으로 낮게 나타났으며, 특히 말리노이즈와 군견 품종에서는 샘플 사이즈가 적어 많은 수의 샘플 사이즈를 이용한 추가 분석이 필요할 것이다.
|
Table 2. Effective population size (Ne) of Army and general dog breeds. 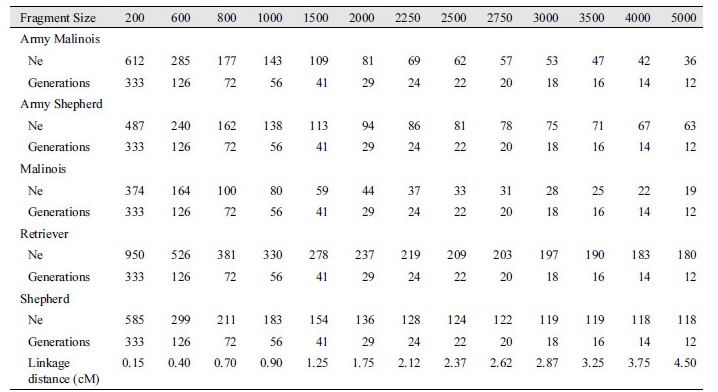
|
|
|
Ne indicates effective population size, and Generation means generation ago. |
|
세계자연보존연맹(IUCN.org)에서는 약 50개체 미만의 유효집단으로 구성된 집단은 종 자체가 심각하게 위협받고 있다고 보고 있다. 이를 근거로 본다면 본 연구에서 분석한 몇몇 종은 심각한 종 위협을 받고 있다. 이러한 결과를 바탕으로 이형 유전자 감소(Heterozygosity Decay) 분석을 수행하였다.
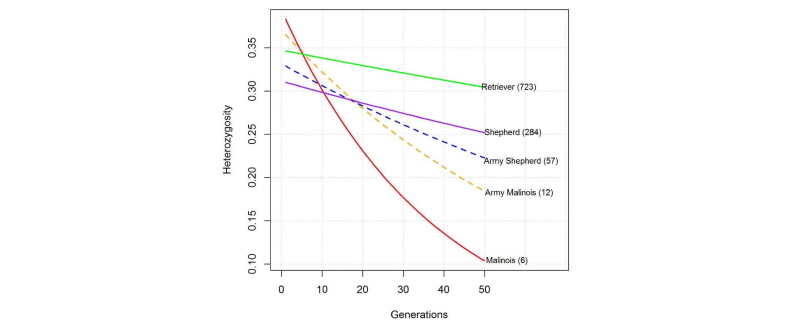
Fig. 4에서 보는 바와 같이 유효 집단의 크기가 18마리였던 일반 집단의 말리노이즈는 약 50세대가 지나서 이형질 대립유전자가 0.1이하로 떨어지는 것으로 분석되었다. 이러한 결과는 약 50세대가 지나서 이 집단은 근친으로 더 이상 집단을 유지할 수 없을 것으로 보인다. 또한 군견 말리노이즈는 유효 집단의 크기가 35두 였는데, 이는 약 50세대가 지나서 이형 대립유전자의 빈도가 0.18로 감소할 것으로 보인다. 현 세대에 이형질 대립유전자가 0.38로 분석되었지만 집단의 수가 급격히 감소함으로 이형대립유전자의 수가 현저히 감소하여 집단의 유전적 다양성이 매우 작아질 것으로 판단된다. 그러나, 일반 리트리버와 세퍼드는 세대가 지속적으로 지나도 이형 대립유전자의 빈도는 크게 바뀌지 않는 것으로 분석되었다. 즉, 일반 집단의 리트리버는 이형대립유전자의 빈도가 0.35에서 0.33으로 감소될 것으로 예측되었으며, 일반 집단 세퍼드의 경우는 현재 이형 대립유전자의 빈도가 0.31이었으나 50세대 이후에 0.27로 감소 될 것으로 예측되었다. 본 연구에서 분석한 결과, 군견 집단에 있어서 말리노이즈와 세퍼드는 집단의 수를 적절히 잘 유지한다면 크게 이형 대립유전자의 빈도가 크게 감소하지 않을 것으로 판단된다. 다만 본 연구에서 분석한 군견 말리노이즈의 집단이 12마리여서 집단을 대표할 수 있는 표본의 크기가 아닐 것으로 판단된다.